Рибосомы. Роль РНК в архитектуре и функциях рибосом
| Загрузить архив: | |
| Файл: ref-28587.zip (43kb [zip], Скачиваний: 76) скачать |
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ
ФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО ОБРАЗОВАНИЮ
НОВОСИБИРСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
<
>
ДОКЛАД
по дисциплине:
«Введение в естествознание»
«Рибосомы. Роль РНК в архитектуре
и функциях рибосом.»
Факультет: ФЕН
Группа: 7403
Студент: Романова Т. Е.
Новосибирск, 2018 г.
План:
Принципы функционирования, роль РНК
Список литературы
1. История исследования рибосомы
Рибосомы впервые были описаны как уплотненные частицы, или гранулы, клеточным биологом румынского происхождения Джорджем Паладе в середине 1950-х годов. В 1974 г. Паладе, Клод и Кристиан Де Дюв получили Нобелевскую премию по физиологии и медицине «за открытия, касающиеся структурной и функциональной организации клетки». Термин "рибосома" был предложен Ричардом Робертсом в 1958 вместо "рибонуклеобелковая частица микросомальной фракции". Биохимические и мутационные исследования рибосомы начиная с 1960-х позволили описать многие функциональные и структурные особенности рибосомы. В начале 2000-х появились атомные структуры отдельных субъединиц, а также полной рибосомы, связанной с различными субстратами, которые позволили понять механизм декодинга (распознавания антикодона тРНК, комплементарного кодону мРНК) и детали взаимодействий между рибосомой, антибиотиками, тРНК и мРНК.
2. Строение и функции, разновидности рибосом
Рибосома - крупный внутриклеточный макромолекулярный ансамбль, ответственный за синтез полипептидной цепи из аминокислот (трансляцию); состоит из молекул РНК (т. наз. рибосомные рибонуклеиновые кислоты, или рРНК) и белков.
Основная масса рибосом локализована в цитоплазме. В бактериальной клетке рибосомы составляют до 30% ее сухой массы: на одну бактериальную клетку приходится примерно 104 рибосом. В эукариотических клетках (клетки всех организмов, за исключением бактерий и синезеленых водорослей) относительное содержание рибосом меньше, и их количество очень сильно варьирует в зависимости от белок-синтезирующей активности соответствующей ткани или отдельной клетки.
В эукариотической клетке все рибосомы цитоплазмы (как мембрано-связанные, так и свободные) образуются в ядрышке; считается, что там они неактивны. Эукариотическая клетка имеет также специальные рибосомы в митохондриях (у животных и растений) и хлоропластах (у растений). Рибосомы этих органелл отличаются от цитоплазматических размерами и некоторыми функциональными свойствами. Они образуются непосредственно в этих органеллах.
Различают два основных типа рибосом. Всем прокариотическим организмам (бактерии и синезеленые водоросли) свойственны так называемые 70S рибосомы, характеризующиеся коэффициентом (константой) седиментации около 70 единиц Сведберга, или 70S (по коэф. седиментации различают и рибосомы других типов, а также субчастицы и биополимеры, входящие в состав Р.). Их молекулярная масса составляет 2,5 · 106, линейные размеры 20-25 нм. По химическому составу это рибонуклеопротеиды; они состоят только из рРНК и белка (соотношение этих компонентов 2:1). Рибосомная РНК в рибосоме присутствует в основном в виде Mg-соли (по-видимому, частично и в виде Са-соли); магния в рибосоме до 2% от сухой массы. Кроме того, в различных количествах (до 2,5%) могут присутствовать также катионы аминов-спермина H2N(CH2)3NH(CH2)4NH(CH2)3NH2, спермидина H2N(CH2)3NH(CH2)4NH2 и др.
Поскольку коэффициенты седиментации зависят не только от молекулярной массы, но и от формы частиц, седиментационные коэффициенты при диссоциации неаддитивны: так, например, бактериальные рибосомы с молекулярной массой ~3*106 Дальтон имеет коэффициент седиментации 70S, обозначается как 70S и диссоциирует на субъединицы 50S и 30S: 70S <
50S + 30S>
Рибосомные субчастицы содержат по одной молекуле рРНК большой длины, масса которой составляет ~1/2 - 2/3 массы рибосомной субчастицы, таким образом, в случае бактериальных рибосом 70S субчастица 50S содержит рРНК 23S (длина ~3000 нуклеотидов) и субчастица 30S содержит рРНК 16S (длина ~1500 нуклеотидов); большая рибосомная субчастица кроме «длинной» рРНК содержит также одну или две «коротких» рРНК (5S рРНК бактериальных рибосомных субчастиц 50S или 5S и 5.8S рРНК больших рибосомных субчастиц эукариот).
Цитоплазма клеток всех эукариотических организмов содержит несколько более крупные 80S рибосомы. Их молекулярная масса около 4·106, линейные размеры 25-30 нм, содержание белка в них значительно больше, чем в прокариотической рибосоме (соотношение РНК: белок ок. 1:1). Рибосомная РНК 80S также связана в основном с Mg и Са и с небольшим кол-вом полиаминов (спермин, спермидин и др.).
Хлоропласты и митохондрии эукариотических клеток содержат рибосомы, отличные от типа 80S. Рибосомы хлоропластов высших растений принадлежат к истинному 70S типу. Митохондриальные рибосомы более разнообразны; их строение находится в зависимости от таксономической принадлежности организма (т.е. от принадлежности к определенному виду, роду или семейству). Напр., митохондриальные рибосомы млекопитающих существенно мельче типичных 70S Р.; коэффициент седиментации этих рибосом составляет около 55S (т. наз. минирибосомы).
Рибосомы из самых разнообразных организмов имеют сходное строение. Они состоят из двух разделяемых субчастиц, или рибосомных субъединиц. При определенных условиях (например, при понижении концентрации Mg2 + в среде) рибосома обратимо диссоциирует на две субчастицы с соотношением их молекулярных масс около 2:1. Прокарйотическая 70S рибосома диссоциирует на субъединицы с коэффициентом седиментации 50S (молекулярная масса 1,5·106) и 30S (молекулярная масса 0,85·106). Эукариотическая рибосома разделяется на субчастицы 60S и 40S. Две рибосомные субчастицы объединены в полную рибосому строго определенным образом, предполагающим специфические контакты их поверхностей.
Как прокариотические, так и эукариотические рибосомы содержат две различные высокомолекулярные рРНК (по одной на каждую субчастицу) и одну относительно низкомолекулярную рРНК в большой субчастице.
Рибосомные белки большинства животных представлены в основном умеренно основными полипептидами, хотя имеется несколько нейтральных и кислых белков. Молекулярные массы рибосомных белков варьирует от 6 тыс. до 60 тыс. г/моль. В прокариотической рибосоме малая субчастица (30S) содержит около 20, большая (50S) - около 30 различных белков; в эукариотической рибосоме 40S субчастица включает около 30 белков, а 60S-около 40 (обычно рибосомы не содержат двух или нескольких одинаковых белков). Рибосомные белки характеризуются глобулярной компактной конформацией с развитой вторичной и третичной структурой; они занимают преимущественно периферическое положение в ядре, состоящем из рРНК. Плотность упаковки рРНК в рибосомах достаточно высока.
По-видимому, рРНК определяет основные структурные и функциональные свойства рибосом, в частности обеспечивает целостность рибосомных субъединиц, обусловливает их форму и ряд структурных особенностей. Специфическая пространственная структура рРНК детерминирует локализацию всех рибосомных белков, играет ведущую роль в организации функциональных центров рибосом.
3. Принципы функционирования, роль РНК
Биосинтез белка имеет два аспекта: химический и генетический. Принципиальным моментом является то, что в природе белок строится из аминокислот не посредством их прямой конденсации с освобождением воды или одновременной полимеризации на матрице, а путем последовательного добавления аминокислотных остатков к одному из концов растущей полипептидной цепи (удлинения) с одновременным сканированием матричного полинуклеотида (мРНК), задающего порядок добавления различных аминокислотных остатков. Три последовательные химические реакции приводят к включению (добавлению) аминокислоты в полипептидную цепь строящегося белка.
Разделение декодирующей и энзиматической функций между субчастицами
Трансляция начинается с того, что мРНК, синтезируемая на ДНК в качестве копии одной из двух цепей последней, связывается с рибосомной частицей. При этом рибосомная частица (у прокариот прямо и непосредственно, а у эукариот после некоторого скольжения вдоль некодирующей части мРНК) специфически взаимодействует с началом кодирующей нуклеотидной последовательности мРНК. Этап связывания мРНК с рибосомной частицей и нескольких последующих событий, приводящих к образованию первой пептидной связи, называется инициацией трансляции. Вслед за инициацией рибосома последовательно "читает" цепочку мРНК по тройкам (триплетам) нуклеотидов по направлению к 3'-концу, наращивая (удлиняя) полипептидную цепочку аминокислотными остатками; этот этап собственно трансляции называется элонгацией. Наконец, достигнув специального нуклеотидного триплета - стоп-кодона, или кодона терминации, - рибосома освобождает синтезированную полипептидную цепочку белка: происходит терминация трансляции.
Генетические функции малой рибосомной субчастицы.
Характерным моментом инициации трансляции является то, что на этом этапе участвуют не целые рибосомы, а их отдельные субчастицы. Другими словами, для того чтобы инициировать трансляцию, рибосома должна быть диссоциирована на две составляющие ее неравные субчастицы. Для этого клетка располагает специальными механизмами, обеспечивающими диссоциацию рибосом после терминации трансляции. Именно малая субчастица рибосомы (30S у прокариот и 40S у эукариот), и только она, связывается с мРНК, то есть служит первичным приемником генетической информации для белоксинтезирующего аппарата. Лишь впоследствии, при завершении этапа инициации трансляции, к ней присоединяется большая субчастица (50S у прокариот и 60S у эукариот), образуя полную рибосомную частицу (70S у прокариот и 80S у эукариот), которая и будет производить элонгацию.
<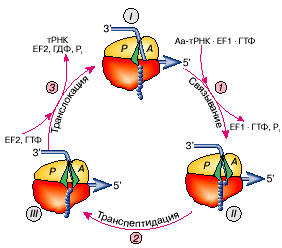
>
Рис. 1: Элементарный элонгационный цикл рибосомы, в результате которого прочитывается один триплет (кодон) мРНК и образуется одна пептидная связь (добавляется одна аминокислота к растущему полипептиду).
В процессе элонгации рибосома удерживает мРНК и движется относительно ее (или протягивает ее сквозь себя) в направлении от 5'-конца к 3'-концу. Удержание мРНК на рибосоме есть целиком и полностью функция малой рибосомной субчастицы, в то время как большая субчастица с мРНК никак не взаимодействует. Соответственно последовательное сканирование кодирующей последовательности мРНК (считывание генетической информации) в ходе элонгации осуществляется на малой субчастице транслирующей рибосомы.
Механизм потриплетного сканирования мРНК в ходе элонгации предполагает участие молекул тРНК, которые взаимодействуют прежде всего с малой рибосомной субчастицей. Малая субчастица в составе полной транслирующей рибосомы имеет два тРНК-связывающих участка, обозначаемых как аминоацил-тРНК-связывающий участок (А-участок) и пептидил-тРНК-связывающий участок (Р-участок). На этапе элонгации Р-участок всегда занят остатком тРНК.
Рассмотрение элементарного акта элонгации удобно начать с момента, когда Р-участок занят молекулой пептидил-тРНК (тРНК, несущая растущую полипептидную цепь), а А-участок вакантен и содержит лишь некий нуклеотидный триплет (кодон) мРНК, пока не взаимодействующий ни с каким триплетом (антикодоном) тРНК (рис. 1, состояние I). Такая рибосома готова (компетентна) связать аминоацил-тРНК, антикодон которой комплементарен триплету (кодону), установленному в А-участке. При наличии около рибосомы такой аминоацил-тРНК происходит первый шаг элементарного элонгационного цикла - кодонспецифическое связывание аминоацил-тРНК с А-участком. Теперь рибосома несет "старую" пептидил-тРНК в Р-участке и новоявленную аминоацил-тРНК в А-участке, которые расположены рядом, бок о бок (рис. 1, состояние II ). Следовательно, в результате кодон-антикодонового взаимодействия мРНК с тРНК на малой субчастице рибосомы произошло декодирование триплета мРНК: именно тот аминокислотный остаток, который был привешен к тРНК с комплементарным антикодоном, оказался в рибосоме.
Далее молекулы пептидил-тРНК и аминоацил-тРНК, расположенные рядом в рибосоме, реагируют друг с другом: пептидильный остаток переносится на аминогруппу молекулы аминоацил-тРНК. Это второй шаг элементарного элонгационного цикла - транспептидация, когда полипептидная цепь удлиняется на одну аминокислоту - на ту, которую принесла тРНК, связавшаяся с А-участком. А сама тРНК, принесшая эту аминокислоту, так и осталась с ней связанной и, таким образом, связанной с удлиненным полипептидом (рис. 1, состояние III). В этом состоянии, однако, новообразованная пептидил-тРНК - точнее, ее остаток тРНК - занимает "не положенный ей" А-участок, а "сидит" в Р-участке деацилированная (без пептидильного или аминоацильного остатков) тРНК. Такое состояние называется претранслокационным. Дальше элонгация идти не может, пока не осуществится третий шаг элонгационного цикла - транслокация, которая выбросит деацилированную тРНК из Р-участка и переведет пептидил-тРНК из А-участка в Р-участок вместе со связанным с ней кодоном мРНК. В результате в освободившемся А-участке на малой рибосомной субчастице установится следующий (новый) кодон мРНК.
Цикл завершился, приведя к образованию одной пептидной связи и соответствующему удлинению растущего полипептида на одну аминокислоту, с одной стороны, и к прочтению одного кодона мРНК и перемещению мРНК на один триплет - с другой. Повторение таких элементарных циклов и создает процесс элонгации.
Таким образом, малая рибосомная субчастица в изолированном состоянии воспринимает копию гена в форме мРНК и инициирует процесс ее трансляции, а в ходе трансляции малая субчастица полной рибосомы удерживает мРНК на себе, декодирует ее с помощью тРНК и последовательно перебирает ее кодоны и тРНК, используя механизм транслокации. Так как все это операции с генетическим материалом, то указанные функции малой рибосомной субчастицы могут быть определены как генетические.
<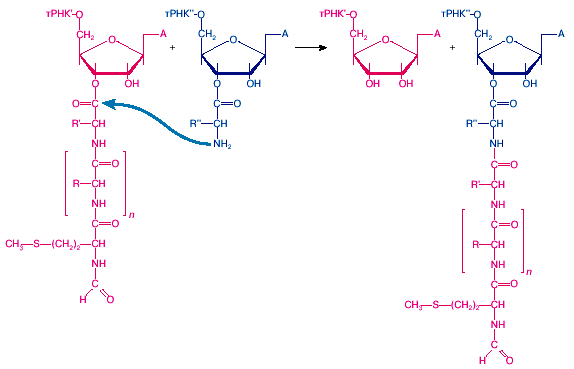
>
Рис. 2: Реакция транспептидации, катализируемая пептидилтрансферазным центром большой рибосомной субчастицы
Энзиматические функции большой рибосомной субчастицы.
Когда пептидил-тРНК занимает Р-участок, а аминоацил-тРНК - А-участок на малой субчастице рибосомы (см. рис. 1, состояние II) , концы остатков тРНК с присоединенными к ним аминоацильными остатками взаимодействуют с большой субчастицей рибосомы. Участок этого взаимодействия на большой субчастице является пептидилтрансферазным центром рибосомы: он катализирует реакцию транспептидации между пептидил-тРНК и аминоацил-тРНК, то есть перенос карбоксильной группы пептидильного остатка на аминогруппу аминоацил-тРНК (рис. 2). В результате образуется новая пептидная связь, и пептидильный остаток становится на одну аминокислоту длиннее. Таким образом, большая субчастица транслирующей рибосомы выступает здесь как фермент, ответственный за образование пептидных связей и в целом за синтез (элонгацию) полипептидной цепи. Это главная энзиматическая функция рибосомы.
Следует отметить, что никакого отдельного от рибосомы белка-фермента, катализирующего образование пептидных связей на рибосоме, не существует. Не найдено и никакого специального белка в составе рибосомы, который бы обладал такой энзиматической функцией. Транспептидация катализируется пептидилтрансферазным центром самой рибосомы как интегральной частью большой рибосомной субчастицы, и основной вклад в организацию центра вносит, по-видимому, рибосомная РНК субчастицы.
Кроме катализа реакции транспептидации большая рибосомная субчастица определенным образом участвует в энзиматическом расщеплении (гидролизе) гаунозинтрифосфата (ГТФ) в процессе трансляции. Дело в том, что, как видно на рис. 1, первый и третий шаги элонгационного цикла идут с участием специальных нерибосомных белков - так называемых факторов элонгации EF1 и EF2. Эти белки являются катализаторами соответствующих нековалентных переходов - связывания аминоацил-тРНК и транслокации. Для такого катализа необходимым оказывается сопряженный гидролиз ГТФ. Именно большая рибосомная субчастица взаимодействует с факторами элонгации и индуцирует гидролиз ГТФ на них. Хотя сам ГТФазный центр находится не на рибосомной субчастице, а на белке - факторе элогнации, ее временная ассоциация с фактором существенна для формирования активного энзиматического ГТФазного центра.
Таким образом, существует четкое разделение труда между двумя неравными субчастицами рибосомы: малая субчастица выполняет генетические функции, будучи ответственной за прием и декодирование генетической информации, в то время как большая участвует в энзиматических реакциях в процессе трансляции.
Конформационная подвижность рибосомы
Работа рибосомы в качестве "лентопротяжного механизма" последовательное прочитывание цепи мРНК от одного конца к другому) в ходе элонгации и ее способность перебрасывать сравнительно большие молекулярные массы (молекулы тРНК) из одного участка в другой в каждом элементарном элонгационном цикле (см. рис. 1, шаг 3 ) предполагают ее механическую подвижность. Взаимная подвижность двух рибосомных субчастиц может быть основным видом крупноблочной подвижности рибосомы в ходе работы, и имеются экспериментальные свидетельства в пользу такой подвижности. Кроме того, существуют указания на подвижность "головки" малой рибосомной субчастицы относительно ее "тела" и на подвижность палочкообразного бокового выступа большой рибосомной субчастицы.
Рибосома при прохождении элонгационного цикла осциллирует между двумя конформационными состояниями: закрытым (сомкнутым) и открытым (разомкнутым). В сомкнутом состоянии рибосомные лиганды (тРНК) зажаты между субчастицами, связаны максимальным количеством контактов с рибосомой и не имеют внутририбосомной подвижности. В разомкнутом состоянии рибосомы лиганды более подвижны, контакты с рибосомой менее полны, и имеется возможность их входа и выхода из рибосомы. Так, на первом этапе связывания аминоацил-тРНК рибосома должна быть открыта для приема лиганда. Возможно, это открытое состояние фиксируется фактором элонгации EF1. Далее EF1 уходит, рибосомные субчастицы плотно смыкаются, и аминоацильный конец связавшейся аминоацил-тРНК вступает в контакт с пептидилтрансферазным центром большой субчастицы. В сомкнутом состоянии пептидил-тРНК и аминоацил-тРНК тесно сближены, и между ними происходит реакция транспептидации. Теперь, чтобы выбросить деацилированную тРНК из рибосомы и дать свободу для перемещения остатка тРНК молекулы пептидил-тРНК из А-участка в Р-участок, рибосому надо приоткрыть, в частности путем раздвигания субчастиц. Это может осуществляться фактором элонгации EF2. После ухода EF2 с рибосомы она снова смыкается и ждет прихода очередной аминоацил-тРНК с фактором элонгации EF1.
Процесс периодического смыкания-размыкания рибосомы является энергозависимым: факторы элонгации EF1 и EF2 взаимодействуют с рибосомой только будучи связанными с ГТФ (согласно модели, при этом взаимодействии происходит открывание рибосомы), а взаимодействие с рибосомой наводит ГТФазную активность, ГТФ гидролизуется, фактор элогации теряет сродство к рибосоме и уходит, и рибосома закрывается. Таким образом, на каждое смыкание-размыкание рибосомы расходуется одна молекула ГТФ. Так как в каждом элонгационном цикле рибосома смыкается-размыкается дважды, то две молекулы ГТФ расходуются на каждый цикл. Это есть энергетическая плата за эффективное (быстрое и надежное) функционирование рибосомы как молекулярной машины.
4. Список литературы
1. Спирин А.С. Принципы структуры рибосом // Соросовский Образовательный Журнал. 1998. N 11. С. 65-70.
2. Спирин А.С. Молекулярная биология: Структура рибосомы и биосинтез белка. М.: Высш. шк., 1986. 300 с.
3. Спирин А.С. О механизме работы рибосомы: Гипотеза смыкания-размыкания субчастиц // Докл. АН СССР. 1968. Т. 179. С. 1467-1470.
6