The effect of light intensity on the amount of chlorophyll in “Cicer arietinum”
| Разделы | Биология |
| Тип | |
| Формат |
|
| Язык | Английский |
| Сдавался/использовался | Ноябрь/2004г., послана в Cardiff на оценивание по программе Международного Бакалавриата (предварительная оценка 7 из 7) |
| Примечание | от автора: Исследовательская работа, на которую был потрачен ровно год |
| Загрузить архив: | |
| Файл: ref-20843.zip (132kb [zip], Скачиваний: 187) скачать |
Extended Essay
Biology (SL)
“The effect of light intensity on the amount of chlorophyll in “Cicer arietinum”
Word count: 4413 words
Content
Abstract ……………………………………………………………………………… 2
Introduction ………………………………………………………………………….. 3
Hypothesis …………………………………………………………………………… 3
Method:
Description ..………………………………………………………………………….. 8
Results ……………………………………………………………………………….. 10
Discussion ……………………………………………………………………..…….. 14
Conclusion ………………………………………………………………………..….. 14
Evaluation of the method ………………………………………………………..…… 15
Bibliography …………………………………………………………………………. 16
Abstract.
Plants, growing on the shaded area has less concentrated green color and are much longer and thinner than plants growing on the sun areas as they are dark green, short and thick. Research question was: “How does the amount of chlorophyll-a and chlorophyll-b, gram per gram of plant, depends on the light intensity in which plants are placed?”
Hypothesis suggests that there are several inner and outer factors that affect the amount of chlorophylls a and b in plants and that with the increase of light intensity the amount of chlorophyll will also increase until light intensity exceeds the value when the amount of destructed chlorophylls is greater than formatted thus decreasing the total amount of chlorophylls in a plant.
The seeds of Cicer arietinum were divided into seven groups and placed into various places with different values of light intensities. Light intensities were measured with digital colorimeter. After three weeks length was measured. Then plants were cut and quickly dried. Their biomass was also measured. Three plants from each group were grinded and the ethanol extract of pigments was prepared. The amount of chlorophylls was measured using method of titration and different formulas.
The investigation showed that plants growing on the lowest light intensity equal 0 lux contained no chlorophyll and had the longest length. The amount of chlorophyll quickly increased and length decreased with the increase of light intensity from 0 lux to 1200 lux. The amount of chlorophyll in plants unpredictably decreased during light intensity equal to 142 lux and than continued increasing and didn’t start decreasing reaching very high value (1200 lux).
The sudden decrease happened due to mighty existence of some inner genetical damages of seeds which prevented them from normal chlorophyll synthesis and predicted decrease didn’t decrease because extremely high light intensity was not exceeded.
Word count: 300 words
I. Introduction.
This theme seemed to be attractive for me because I could see that results of my investigation could find application in real life.
While walking in the forest in summer I saw lots of plants of different shades of green color: some of them were dark green, some were light green and some even very-very light green with yellow shades, hence I became very interested in this situation and wanted to know why it happens to be so. I also saw that those plants that were growing on sunny parts of forest, where trees were not very high, had dark green color and those, that were growing in shady parts of the same forest had very light green color. They also had difference in their length and thickness – those, that were growing on light were very short, but thick and strong, and those, growing in shady regions were very thin and fragile.
Hence I became very interested in finding scientifical description ofmy observations.
The aim of my project is to find out how does the changes in light intensity affect balance of chlorophyll in Cicer arietinum.
II. Hypothesis.
There are several factors that affect the development of chlorophyll in plants.[1]
Inner factors. The most important one is – genetical potential of a plant, because sometimes happen mutations that follow in inability of chlorophyll formation. But most of the times it happens that the process of chlorophyll synthesis is broken only partly, revealing in absence of chlorophyll only in several parts of the plant or in general low rate of chlorophyll. Therefore plants obtain yellowishcolor. Lots of genes participate in the process of chlorophyll synthesis, therefore different anomalies are widely represented. Development of chloroplasts depends on nuclear and plastid DNA and also on cytoplasmatic and chloroplastic ribosomes.
Full provision of carbohydrates seem to be essential for chlorophyll formation, and those plants that suffer from deficit of soluble carbohydrates may not become green even if all other conditions are perfect. Such leaves, placed into sugar solution normally start to form chlorophyll. Very often it happens that different viruses prevent chlorophyll formation, causing yellow color of leaves.
Outside factors. The most important outside factors, affecting the formation of chlorophyll are: light intensity, temperature, pH of soil, provision of minerals, water and oxygen. Synthesis of chlorophyll is very sensitive to all the factors, disturbing metabolic processes in plants.
Light. Light is very important for the chlorophyll formation, though some plants are able to produce chlorophyll in absolute darkness. Relatively low light intensity is rather effective for initialization and speeding of chlorophyll development. Green plants grown in darkness have yellow color and contain protochlorophyll – predecessor of chlorophyll а, which needs lite to restore until chlorophyll а. Very high light intensity causes the destruction of chlorophyll. Hence chlorophyll is synthesized and destructed both at the same time. In the condition of very high light intensity balance is set during lower chlorophyll concentration, than in condition of low light intensity.
Temperature. Chlorophyll synthesis seems to happen during rather broad temperature intervals. Lots of plants ofумеренной зоны synthesize chlorophyll from very low temperatures till very high temperatures in the mid of the summer. Many pine trees loose some chlorophyll during winters and therefore loose some of their green color. It may happen because the destruction of chlorophyll exceeds its formation during very low temperatures.
Provision with minerals. One of the most common reason for shortage of chlorophyll is absence of some important chemical elements. Shortage of nitrogen is the most common reason for lack of chlorophyll in old leaves. Another one is shortage of ferrum, mostly in young leaves and plants. And ferrum is important element for chlorophyll synthesis. And magnesium is a component of chlorophyll therefore its shortage causes lack of production of chlorophyll.
Water.Relatively low water stress lowers speed of chlorophyll synthesis and high dehydration of plants tissues not only disturbs synthesis of chlorophyll, but even causes destruction of already existing molecules.
Oxygen. With the absence of oxygen plants do not produce chlorophyll even on high light intensity.This shows that aerobic respiration is essential for chlorophyll synthesis.
 Chlorophyll.[2] The synthesis of chlorophyll is induced by
light. With light, a gene can be transcripted and translated in a protein.
Chlorophyll.[2] The synthesis of chlorophyll is induced by
light. With light, a gene can be transcripted and translated in a protein.
The plants are naturally blocked in the
conversion of protochlorophyllide to chlorophyllide. In normal plants these
results in accumulation of a small amount of protochlorophyllide which is
attached to holochrome protein. In vivo at least two types of
protochlorophyllide holochrome are present. One, absorbing maximally at
approximately 650 nm, is immediately convertible to chlorophyllide on exposure
to light. If

 |
Protochlorophyllide
|
POR |
 |
Chlophyllide
 |

 Chlorophyll b Chlorophyll a
Chlorophyll b Chlorophyll a
Chlorophyll[3] is a green compound found in leaves and green stems of plants. Initially, it was assumed that chlorophyll was a single compound but in 1864 Stokes showed by spectroscopy that chlorophyll was a mixture. If dried leaves are powdered and digested with ethanol, after concentration of the solvent, 'crystalline' chlorophyll is obtained, but if ether or aqueous acetone is used instead of ethanol, the product is 'amorphous' chlorophyll.
In 1912, Willstatter et al. () showed that chlorophyll was a mixture of two compounds, chlorophyll-a and chlorophyll-b:

Chlorophyll-a (C55H72MgN4O5, mol. wt.: 893.49). The methyl group marked with an asterisk is replaced by an aldehyde in chlorophyll-b (C55H70MgN4O6, mol. wt.: 906.51).
The two components were separated by shaking a light petroleum solution of chlorophyll with aqueous methanol: chlorophyll-a remains in the light petroleum but chlorophyll-b is transferred into the aqueous methanol. Cholorophyll-a is a bluish-black solid and cholorophyll-b is a dark green solid, both giving a green solution in organic solutions. In natural chlorophyll there is a ratio of 3 to 1 (of a to b) of the two components.
The intense green colour of chlorophyll is due to its strong absorbencies in the red and blue regions of the spectrum, shown in fig. 1. () Because of these absorbencies the light it reflects and transmits appears green.

Fig. 1 - The uv/visible adsorption spectrum for chlorophyll.
Due to the green colour of chlorophyll, it has many uses as dyes and pigments. It is used in colouring soaps, oils, waxes and confectionary.
Chlorophyll's most important use, however, is in nature, in photosynthesis. It is capable of channelling the energy of sunlight into chemical energy through the process of photosynthesis. In this process the energy absorbed by chlorophyll transforms carbon dioxide and water into carbohydrates and oxygen:
CO2
+ H2O  (CH2O) + O2
(CH2O) + O2
Note:
CH2O is the empirical formula of carbohydrates.
The chemical energy stored by photosynthesis in carbohydrates drives biochemical reactions in nearly all living organisms.
In the photosynthetic reaction electrons are transferred from water to carbon dioxide, that is carbon dioxide is reduced by water. Chlorophyll assists this transfer as when chlorophyll absorbs light energy, an electron in chlorophyll is excited from a lower energy state to a higher energy state. In this higher energy state, this electron is more readily transferred to another molecule. This starts a chain of electron-transfer steps, which ends with an electron being transferred to carbon dioxide. Meanwhile, the chlorophyll which gave up an electron can accept an electron from another molecule. This is the end of a process which starts with the removal of an electron from water. Thus, chlorophyll is at the centre of the photosynthetic oxidation-reduction reaction between carbon dioxide and water.
Treatment of cholorophyll-a with acid removes the magnesium ion replacing it with two hydrogen atoms giving an olive-brown solid, phaeophytin-a. Hydrolysis of this (reverse of esterification) splits off phytol and gives phaeophorbide-a. Similar compounds are obtained if chlorophyll-b is used.

Chlorophyll can also be reacted with a base which yields a series of phyllins, magnesium porphyrin compounds. Treatment of phyllins with acid gives porphyrins.

Now knowing all these factors affecting the synthesis and destruction of chlorophyll I propose that the amount of chlorophyll in plant depends on light intensity in the following way: with the increase of light intensity the amount of chlorophyll increases, but then it starts decreasing because light intensity exceed the point when there is more chlorophyll destructed than formed.

|
|

|






|
|
 SHAPE* MERGEFORMAT
SHAPE* MERGEFORMAT 
Variables.
Independent:
- Light intensity, lux
Constant:
- pH of soil
- water supply, ml
- temperature, to C
Dependent:
- length, cm
- amount of chlorophyll in gram of a plant, gram per gram
III. Method.
Apparatus:
· seeds of Cicer arietinum
· 28 plastic pots
· water
· scissors
· ruler (20 cm ± 0.05 cm)
· CaCO3
· soil (adopted for home plants)
· digital luxmeter (± 0.05 lux)
· test tubes
· H2SO4 (0.01 M solution)
· Pipette (5 cm3 ± 0.05 cm3)
· mortar and pestle
· burette
· ethanol (C2H5OH), 98%
· beakers
Firstly I went to the shop and bought germinated seeds of Cicer arietinum. Then sorted seeds and chose the strongest ones. After that I prepared soil for them and put them in it.
As the
aim of this project is to investigate the dependence of mass of chlorophyll in
plants during different light intensities it was needed to create those various
conditions. Pots with seeds were placed into the following places: in the
wardrobe with doors (light intensity is o lux), under the sink (light intensity
is 20,5 lux), in the shell of bookcase (light intensity is 27,5 lux), above the
bookcase (light intensity is 89,5 lux), above the extractor (light intensity is
142 lux), beyond the curtains (light intensity is 680 lux) and on the open sun
(light intensity is 1220 lux). Light intensity was measured with the help of
digital luxmeter.It was measured four times
each day: morning,
Growing plants were provided with the same amount of water (15 ml, once a day in the morning) and they were situated in the same room temperature (20o C), pH of soil was definitely the same because all the plants were put in the same soil (special soil for room flowers).
After three weeks past the length of plants was measured with the help of ruler. Firstly the plants were not cut, so their length had to be measured while they were in the pots. The ruler was placed into the pot and plants were carefully stretched on it. The action was repeated three times and only maximum value was taken into consideration. After that plants were cut. Then those already cut plants were put into the dark place and quickly dried.
Titration.
I have chosen three plants from each light intensity group and measured their weight. . In order to obtain the pigments, three plants were cut into small pieces and placed in a mortar. Calcium carbonate was then added, together with a little ethanol (2 cm3). The leaf was grinded using a pestle until no large pieces of leaf tissue were left, and the remaining ethanol was poured into the mortar (3 cm3). Then 1 ml of obtained solution was placed into the test tube and this 1 ml of solution was then titrated against 0.01 M solution of sulfuric acid, through the use of a pipette. The titration was complete when the green solution turned dark olive-green[4]. This solution obtained from the first action was stored as the etalon for the following ones. The settled olive-green coloring meant that all chlorophyll had reacted with H2SO4. So the process of titration was repeated 7 times for all light intensity groups.
The solution is titrated until the dark olive-green color because it is known that when the reaction between chlorophyll and sulfuric acid happens, chlorophyll turns into phaeophetin which has grey color (see table 1), therefore when the solution is olive-green, than the reaction has succeeded. But while searching in the internet and books I found out that there are several opinions about the color of phaeophytin – in the book written by Viktorov it is ssaid to have grey color, but in the internet link it is said to have brown olive-green color. Also I made chromatography in order to investigate the color of phaeophytin and the result was that it has grey color. It can be proposed that olive-green color is obtained because grey phaeophetyn is mixed with other plant pigments.
So titration is one of the visual methods that can be used in order to find the mass of chlorophyll in plants.
All the measurements and even chromatography were done three times and the mean value was taken, for chromatography grey color was confirmed.
Table 1. Plant pigments.
|
Name of the pigment |
Color of the pigment |
|
Chlorophylls ( a and b ) |
Green |
|
Carotene |
|
|
Xanitophyll |
Yellow |
|
Phaeophytin-a |
OLIVE BROUN or GREY |
IV. Results.
Table 2. Raw data.
|
Number of plant |
Light intensity (lux) |
||||||
|
0,0 |
20,5 |
27,5 |
89,5 |
142,0 |
680,0 |
1220,0 |
|
|
1 |
23 |
35 |
20 |
1 |
30 |
2 |
15 |
|
2 |
30 |
36 |
33 |
4 |
31 |
20 |
16 |
|
3 |
38 |
37 |
35 |
8 |
34 |
21 |
16 |
|
4 |
39 |
37 |
36 |
9 |
35 |
21 |
16 |
|
5 |
44 |
38 |
37 |
9 |
38 |
21 |
17 |
|
6 |
46 |
39 |
40 |
12 |
38 |
22 |
17 |
|
7 |
50 |
39 |
40 |
12 |
38 |
22 |
19 |
|
8 |
52 |
40 |
43 |
13 |
39 |
23 |
20 |
|
9 |
55 |
40 |
43 |
15 |
39 |
25 |
21 |
|
10 |
40 |
18 |
40 |
27 |
22 |
||
|
11 |
42 |
20 |
41 |
29 |
26 |
||
|
12 |
42 |
22 |
41 |
30 |
|||
|
13 |
42 |
22 |
41 |
31 |
|||
|
14 |
42 |
24 |
42 |
33 |
|||
|
15 |
43 |
25 |
42 |
34 |
|||
|
16 |
43 |
25 |
43 |
34 |
|||
|
17 |
44 |
25 |
43 |
35 |
|||
|
18 |
44 |
25 |
43 |
35 |
|||
|
19 |
45 |
26 |
45 |
37 |
|||
|
20 |
45 |
26 |
45 |
38 |
|||
|
21 |
45 |
26 |
46 |
38 |
|||
|
22 |
45 |
26 |
46 |
41 |
|||
|
23 |
46 |
27 |
48 |
41 |
|||
|
24 |
46 |
29 |
48 |
44 |
|||
|
25 |
49 |
32 |
49 |
||||
|
26 |
34 |
49 |
|||||
|
Mean |
41,888889 |
41,76 |
36,33333 |
19,80769 |
41,30769 |
29,33333 |
18,63636 |
|
Median |
44 |
42 |
37 |
23 |
41,5 |
30,5 |
17 |
|
St. deviation |
10,50529 |
2,928 |
4,740741 |
7,467456 |
4 |
7,472222 |
2,694215 |
Table 3. Frequency of lengths of 3-weeks-old plants depending on light intensity.
|
Light intensity, lux |
|||||||
|
Plant length, cm (class) |
0,0 |
20,5 |
27,5 |
89,5 |
142,0 |
680,0 |
1220,0 |
|
0.0-10.0 |
0 |
0 |
0 |
5 |
0 |
1 |
0 |
|
10.1-20.0 |
0 |
0 |
1 |
6 |
0 |
1 |
8 |
|
20.1-30.0 |
2 |
0 |
0 |
13 |
1 |
10 |
3 |
|
30.1-40.0 |
2 |
9 |
6 |
2 |
9 |
9 |
0 |
|
40.1-50.0 |
3 |
15 |
2 |
0 |
16 |
3 |
0 |
|
50.1-60.0 |
2 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Total |
9 |
24 |
9 |
26 |
26 |
24 |
11 |
Table 3 (alternative) Frequency of length of 3-weeks-old plants depending on light intensity.
|
Light intensity, lux |
|||||||
|
Plant length (Class) |
0,0 |
20,5 |
27,5 |
89,5 |
142,0 |
680,0 |
1220,0 |
|
0.0-10.0 |
0 |
0 |
0 |
19,23% |
0 |
4,17% |
0 |
|
10.1-20.0 |
0 |
0 |
11,10% |
23,08% |
0 |
4,17% |
72,72% |
|
20.1-30.0 |
0 |
0 |
0 |
50% |
3,85% |
41,62% |
27,28% |
|
30.1-40.0 |
0 |
37,50% |
66,60% |
7,69% |
34,62% |
37,52% |
0 |
|
40.1-50.0 |
0 |
62,50% |
22,30% |
0 |
61,53% |
12,52% |
0 |
|
50.1-60.0 |
100% |
0 |
0 |
0 |
0 |
0 |
0 |
|
Total |
1 |
1 |
1 |
1 |
1 |
1 |
1 |

Calculation of the mean length of plants.
For light intensity equal to20,50 lux:
The sum of lengths of all plants in this group is 45cm + 37cm + 39cm + 49cm + 46cm + 44cm + 45cm + 44cm + 42cm + 37cm + 40cm + 40cm + 39cm + 43cm + 42cm + 42cm + 36cm + 45cm + 38cm + 45cm + 46cm + 40cm + 35cm + 42cm + 43cm = 1044cm
Hence mean length is 1044cm : 25 plants = 41,76cm
Table 4.
|
Light intensity, lux |
Mean wet biomass, g |
Mean dry biomass, g |
% of water |
Mean length, cm |
Mass of chl. In 1 g |
|
0 |
0,273 |
0,041 |
84,98 |
41,89 |
0,0000 |
|
20,5 |
0,579 |
0,056 |
90,33 |
41,76 |
0,0496 |
|
27,5 |
0,332 |
0,033 |
90,06 |
36,33 |
0,1462 |
|
89,5 |
0,181 |
0,018 |
90,06 |
19,81 |
0,1769 |
|
142 |
0,511 |
0,047 |
90,80 |
41,33 |
0,0697 |
|
680 |
0,338 |
0,043 |
87,28 |
29,33 |
0,1557 |
|
1220 |
0,301 |
0,034 |
88,70 |
18,64 |
0,1939 |
Calculation of amount of chlorophyll in plants basing on the results of titration
H2 SO4+ C56 O5 N4 Mg => C56 O5 N4 H + MgSO4
Concentration of H2SO4 is 0,01 M
C – concentration
V – volume
n – quantity of substancy
m – mass
Mr – molar mass
For light intensity equal to 20,5 lux.
n = V (in dm3) ∙ C
2 ∙ 10-3 ∙ 0,01 = 2 ∙ 10-5
n = m / Mr => m = n ∙ Mr
m = 2 ∙ 10-5 ∙ 832 = 1,664 ∙ 10-2 grams
mass of plant mass of chlorophyll
1,68 grams - 0,08335 grams of chlorophyll
1 gram - x grams of chlorophyll
Hence there are 0,0496 grams of chlorophyll.

Table 5. The correlation between mean length of plants and mean dry biomass.
|
Site |
Mean length, cm |
Rank (R1) |
Mean dry biomass, g |
Rank (R2) |
D (R1-R2) |
D2 |
|
|
1 |
41,89 |
1 |
0,041 |
4 |
-3 |
9 |
|
|
2 |
41,76 |
2 |
0,056 |
1 |
1 |
1 |
|
|
3 |
36,33 |
4 |
0,033 |
6 |
-2 |
4 |
|
|
4 |
19,81 |
6 |
0,018 |
7 |
-1 |
1 |
|
|
5 |
41,33 |
3 |
0,047 |
2 |
1 |
1 |
|
|
6 |
29,33 |
5 |
0,043 |
3 |
2 |
4 |
|
|
7 |
18,64 |
7 |
0,034 |
5 |
2 |
4 |
|
|
Rs = 0,57 |
|||||||
|
criticalvalue = 0,79 |
|||||||
|
|
|
0,57<0,79, therefore there is no significant correlation between mean length of plants and mean dry biomass. |


Table 6. The correlation between mean length and mass of chlorophyll per 1 g of plant.
|
Site |
Mean length, cm |
Rank (R1) |
Mass of chl. In 1 g |
Rank (R2) |
D (R1-R2) |
D^2 |
|
|
1 |
41,89 |
1 |
0,0000 |
7 |
-6 |
36 |
|
|
2 |
41,76 |
2 |
0,0496 |
6 |
-4 |
16 |
|
|
3 |
36,33 |
4 |
0,1462 |
4 |
0 |
0 |
|
|
4 |
19,81 |
6 |
0,1769 |
2 |
4 |
16 |
|
|
5 |
41,33 |
3 |
0,0697 |
5 |
-2 |
4 |
|
|
6 |
29,33 |
5 |
0,1557 |
3 |
2 |
4 |
|
|
7 |
18,64 |
7 |
0,1939 |
1 |
6 |
36 |
|
|
|||||||
|
There is negative correlation between mean length of plants and mass of chlorophyll per 1 g of plant |


Table 7. The correlation between mean dry biomass and mass of chlorophyll per 1 g of plant.
|
Site |
Mean dry biomass, g |
Rank (R1) |
Mass of chl. In 1 g |
Rank (R2) |
D (R1-R2) |
D^2 |
|
|
1 |
0,041 |
4 |
0,0000 |
7 |
-3 |
9 |
|
|
2 |
0,056 |
1 |
0,0496 |
6 |
-5 |
25 |
|
|
3 |
0,033 |
6 |
0,1462 |
4 |
2 |
4 |
|
|
4 |
0,018 |
7 |
0,1769 |
2 |
5 |
25 |
|
|
5 |
0,047 |
2 |
0,0697 |
5 |
-3 |
9 |
|
|
6 |
0,043 |
3 |
0,1557 |
3 |
0 |
0 |
|
|
7 |
0,034 |
5 |
0,1939 |
1 |
4 |
16 |
|
|
Rs = -0,57 |
|||||||
|
|
|||||||
|
0,57<0,79, therefore there is no significant correlation between mean dry biomass and mass of chlorophyll per 1 g of plant |


V. Discussion.
Several tendencies can be clearly seen.
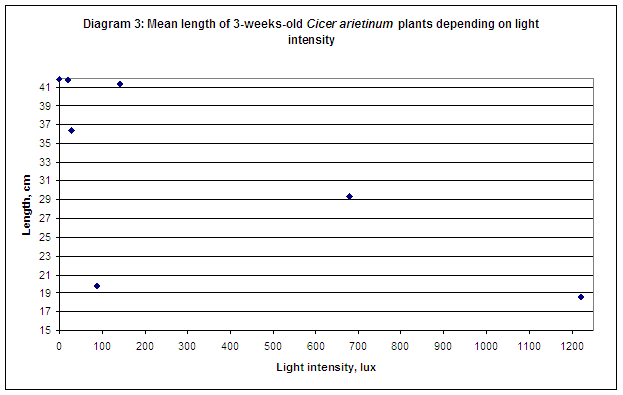
For the first, with the increase of light intensity mean length of plants is decreasing, but there are exceptions. For light intensity 142 lux the value of mean length is approximately equal to the values of length for light intensities 0 lux and 20,5 lux. If exclude this data it is also seen that for light intensity equal to 680 lux mean length is also slightly falling out from the main tendency – decreasing from 19.81 cm.
The second tendency is increase of mass of chlorophyll per 1 gram of plant biomass with the increase of light intensity. But the values of mass of chlorophyll of those plants under light intensities 142 lux and 680 lux are falling out from the main tendency. The first and the second ones are too small – approximately equal to the value corresponding to 20.5 lux light intensity and to 89.5 lux respectively. This may happen because not all the seeds of Cicer arietnum were of the same quality, because it is impossible to guarantee that more than 250 seeds in one box have the same high quality. At the mean time it was expected that starting from the light intensity more than 680 lux the amount of chlorophyll in plants will decrease, because the value of destructed chlorophyll with be bigger than the value of newly formatted. But the experiments showed that the amount of chlorophyll was constantly increasing even when the light intensity level exceeded the point 1220 lux. This could happen because light intensity equal to 1220 lux is not so extremely high that the amount of total chlorophyll in plants will start decreasing.
Also it is clearly seen that there are no correlations between light intensity and values of wet and dry biomass.
Basing on these arguments the sudden decrease of the amount of chlorophyll in plants placed on light intensity equal to 142 lux was likely to be insignificant and could not be considered as a trend.
But it is impossible to forget such important factor as plant hormones that affect the growth and development of plants. There are five generally accepted types of hormones that influence plant growth and development. They are: auxin, cytokinin, gibberellins, abscic acid, and ethylene. It is not one hormone that directly influences by sheer quantity. The balance and ratios of hormones present is what helps to influence plant reactions. The hormonal balance possibly regulates enzymatic reactions in the plant by amplifying them.
VI. Conclusion.
Due to results of my investigation it is seen that my hypothesis didn’t confirm fully (for example, comparing the diagram 1 and diagram 7), because I proposed that when light intensities will be very high, mass of chlorophyll in plant will start decreasing and due to my observations it didn’t happen. I should say that the only reason I can suggest is that I haven’t investigated such extremely high light intensities, so that chlorophyll start destructing. But if we will not pay attention to that fact the other part of my hypothesis was confirmed and mass of chlorophyll in plants increased with the increase of light intensity. Furthermore I didn’t estimate amount of plant hormones and so didn’t estimate their influence on results.
Questions for further investigation:
1.Investigating very high light intensities.
2.Implementation of colorimetric analysis.
3.Paying attention to estimation of plant hormones level.
Those questions should be further investigated in order to get clearer picture and more accurate results of the dependence of the amount of chlorophyll in plants on the light intensity, knowing the fact that the amount of chlorophyll has a tendency to decrease at extremely high light intensities. So this statement needs an experimental confirmation and as in this investigation conditions with extremely light intensity were not created in further investigations they have to be created.
Implementation of colorimetric analysis is
also very important thing, because it gives much more accurate results
comparing with the titration method. The colorimetric method suggests that as different pigments absorb
different parts of light spectrum differently, the absorbance of a pigments
mixture is a sum of individual absorption spectra. Therefore the quantity of
each individual pigment in a mixture can becalculated using absorbance of the certain colors and molecular
coefficients of each pigment. This was proposed by D.
A. Sims, and J. A. Gamon (
VII. Evaluation.
There are several results in my work, that are falling out from the main tendencies. It may seem that such results may occur due to different percentage of water in plants, but when I was calculating mass of chlorophyll in 1 gram of plant I was using only values of mean dry biomass so it couldn’t affect my results. (see table 3)
At the same time such differences in the percentage of water are easily explained. The rate of evaporation of water from plants, which were put under 1220 lux light intensity was much higher than of those put under 20.5 lux light intensity, therefore percentage of water in the soil may vary, though I provided all the plants with the same volume of water at the same periods of time.
One more reason that could be proposed is the reason connected with the pH of water with which flowers were provided. It was not measured but the thing that could have happened is that it had somehow changed the pH of soil in which seeds were placed and therefore changed the amount of synthesized chlorophyll.
Titration is not a perfect way of obtaining results. This happens because the method is based on visual abilities of a person – he has to decide whether the color he obtained is dark olive-green or not so dark olive-green. Such a situation concerns lots of mistakes due to different optical abilities of each person, even some humans are not able to distinguish those colors, because of the disease called Daltonism.
Even those who do not suffer from this disease can also make mistakes in such experiment. It is known that people who suffer from Myopia can hardly see objects that are far from them, but don’t have problems with objects that are near, but it is also important to take into consideration the fact that their ability to distinguish colors is also lower comparing with humans with normal eyesight.
There also exist the so called human factor, which also affects the investigation. Man can’t be absolutely objective, because sometimes it is too hard for a person to falsify his own theory or hypothesis, so one can ignore results that are not suitable for his statements and select only those that are suitable, which will also affect the investigation not in good way.
So as human’s eye is not a perfect instrument and humans are not perfectly objective there should be other methods of investigating the amount of chlorophyll in plant.
Moreover titration method doesn’t distinguish between chlorophylls-a and chlorophyll-b, phaeophytin-a and phaeophytin-b, as their colors differ, this giving not very accurate results. Also due to this limiting factor it is impossible to know whether the whole amount of chlorophyll reacted with the sulfuric acid and again it adds an uncertainty to the results. Furthermore the saturation of color depends on the extent of dilution and it is nearly impossible to say if the solution was diluted till the same color or not, because it is very difficult to distinguish between different shades of olive green color.
BIBLIOGRAPHY
1) Allott, Biology for IB diploma (standard and higher level), Oxford University Press, ISBN 0-19914818
2) M. Roberts, M. Reisse, G. Monger, Biology: principles and approaches, Nelson, ISBN 0-17-44-8176-4
3) T. King, M. Reiss, M. Roberts, Practical advanced biology, Nelson Thorns, ISBN 0-170448308-
4) Викторов Д. П., Практикум по физиологии растений. – 2-е изд. – Воронеж: ВГУ, 1991
5)
,
6) 05/05/2004
7)
,
8)
,
9)
.
10)
,
11)
12)
,
[1]
[4] Викторов Д. П., Практикум по физиологии растений. – 2-е изд. – Воронеж: ВГУ, 1991, p.66
[5] 05/05/2004