Происхождение эукариотических клеток
| Загрузить архив: | |
| Файл: hai-0286.zip (456kb [zip], Скачиваний: 70) скачать |
КУРСОВАЯ РАБОТА
по биологии
тема:
“Происхождениеэукариотических
клеток”
Выполнила: СуроваВера
10-В класс,школа N55
Проверила: СтруковаН.А.
г.Рязань 1999 год
ПЛАН
1 . Симбиотическая теория:
1.1Фундаментальноеразграничение между прокариотамииэукариотами;
1.2Прокариотические организмы;
1.3Эукариотические клетки;
1.4Митохондрии ипластиды;
1.5Способность эукариоткфотосинтезу;
2.Предшественники жизни;
3. Ферментирующие организмы:
3.1Источники энергиираннихклеток;
3.2Метаногенез какисточникэнергии;
3.3Цианобактерии ихлорооксибактерии;
4.Фотосинтези воздух;
5.Ассоциациии эукариоты:
5.1Теориипоследовательныхсимбиозов;
5.2Симбиотическоеприобретение подвижности;
5.3Эукариоты – фотосинтетики;
6.Взглядна эволюциюклетки:
6.1Гипотеза прямойфилиации;
6.2Гипотеза: от цианобактерийкводорослям;
6.3Совместимость 2 –х гипотез:прямой филиациии
эндосимбиоза;
7.Выводы;
8.Литература .
ТЕОРИЯ ЭНДОСИМБИОЗА
Все живыеорганизмы на Землеобычноделят на прокари-отиэукариот . Главной особенностьюпрокариот вотличиеот эукариотявляетсяотсутствие унихполноценного клеточногоядра , покрытогомембраной . Клеткипрокариотимеют оченьне-большиеразмеры порядка1 мкм .Объем эукариотических клеток, содержащих полноценноеядро,в800 – 1000разбольше объемаклетокпрокариот . Всвязис этим клеткам – эукариотамнеобходим длявыполненияжизненных функцийгораздобольший мембранныйаппарат . Действительно , в эукариотической клеткевыявленысубклеточные структуры , окруженныенезави-симымиот плазмалеммыидруг отдругамембранами . Этиструктурыназываются органеллами . Каждой органелле:митохондрии , хлоропласту ,ядру , лизосомам , аппаратуГольджиит. д .присущисвои специфическиефункциив жизниклетки .
Спомощью молекулярно – биологических подходовполученыубедительные доказательстватого ,чтовсе живыесуществадолжны бытьразделенына 3надцарства: бактерии , археииэукариоты . Бактериииархеи возниклинаЗемле отобщегопредка - такназываемогопрогенота - около4 млрд летназад , аэукариотыпо даннымпалеонтологов появились наЗемлечерез 500 млнлетпосле прокариот .Наиболеепопулярной идостаточнообоснованной идеейпроисхожденияэукариотическихклеток, внастоящеевремя являктсяидеямногократного эндосимбиозаразличных прокариот . Попредставлениямизвестных микробиологовА. КлюйвераиК. Ван Ниля ,именноу древнихпрокариот произошелотборклеток снаиболееэкономичными изачастуюполифункциональнымибиохи-мическимиреакциями , ставшимиосновойважнейших путейметаболизма .
Симбиотическая теория происхождения иэволюции клетокоснована на двух концепциях , новыхдлябиологии .Согласнопервойиз этихконцепций, самое фундаментальное разграни-чениевживой природе- эторазграничение междупрокариотамии эукариотами ,т.е. междубактериями иорганизмами,состоящимииз клетоксистинными ядрами- протистами , животными , грибамиирастениями .Втораяконцепция состоитвтом , чтоисточником некоторых частей эукариотическихклетокбыла эволюциясимбиозов , т.е. формированиепостоянныхассоциаций междуорганизмамиразных видов .Предполагается , чтотрикласса органелл- митохондрии , реснички ифотосинтезирующиепластиды – произошли от свободноживущихбактерий , которыев результатесимбиоза были вопределеннойпоследовательности включены всоставклеток прокариот -хозяев.Эта теориявбольшой мереопираетсяна неодарвинистские представления,развитые генетиками, экологами, цитологамиидругими учеными,которыесвязали Менделевскуюгенетику сдар-виновской идеейестественногоотбора .Онаопираетсятакже насовершенноновые илинедавновозрожденные научныенаправ-ления :на молекулярнуюбиологию ,особенно наданныео структуребелкови последовательностиаминокислот ,намикро палеонтологию , изучающуюнаиболееранние следыжизнина Земле ,и даженафизику ихимиюатмосферы , посколькуэтинауки имеютотношениек газамбиологическогопроисхсждения . Всеорганизмы, состоящиеизклеток, могутбытьсгруппиро-ваныв пять царств :царствопрокариот (Monera,кудаотносятся бактерии )ичетыре царства эукариот (Protoctista, Animalia, Fungi ,Plantae). Протоктисты-этоэукариотическиеорганизмы,неотносящиеся кживотным,грибамили растениям.В царствопротоктистоввходят водоросли ,протозои, слизевики идругие эукариотические организмы неясной принадлежности . Протисты определены более ограничительно-как однокле-точные эукариоты .Такимобразом , царствоProtoctistaвключает нетолькопротистов – одноклеточныхэукариот ,нои ихближай-ших многоклеточныхпотомков,такихкак красныеибурые водоросли ,атакже многиемикроорганизмы ,сходныесгрибами ,напримерхитридиевые . Согласнотрадиционному представ-лению о прямой филиации ,такие клеточныеорганеллы ,какмитохондрии ипластиды , возникли путемкомпартментализациисамой клетки .
НЕСКОЛЬКО ПРОКАРИОТ ОБРАЗУЮТ
ОДНУ ЭУКАРИОТУ
Клетки -этоограниченные мембранами системы , наименьшиеизбиологических единиц ,способных к саморепродукции . Даже мельчайшие клетки содержат генывформе молекулДНК (однойили нескольких )ибелоксинтезирующийаппарат , состоящийизнескольких типовРНКи множества белков.Все клеткисодержатрибосомы – тельцадиаметром около0,02мкм,состоящие изРНКпо меньшеймеретрех типов , и примерно пятидесяти различныхбелков. Прокариотичесие организмы – наименьшие биологические единицы ,которые удовлетворяют этому определениюклетки .Уних нетядер . Сдругойстороны ,одиночные эукариотические( имеющие ядро)клетки в известномсмысле вообщенеявляются единицами . Это комплексы различных белоксинтезирующих единиц ( ну-клеоцитоплазмы ,митохондрий ипластид ) ,типи числокоторых варьирует в зависимостиот видовойпринадлежности . Такимобразом , согласно этой теории , эукариотические клетки возникли в результате кооперации первоначальнонезависимых элементов,объединившихся в определенном порядке. Тэйлор назвал такоепредставление о раздельном происхождении иразвитии частей эукариотической клетки иих последующем объединении- теорией последовательных эндосимбиозов.Условия высокой температуры и кислотности привели к выработке в нуклеоцитоплазме особого классабелков -гистонов - для защиты ее ДНК .Митохондрии были способны расщеплять трехуглеродныесоединения до углекислоты иводы . Симбиотическая ассоци-ация нуклеоцитоплазмы и митохондрий начала изменяться всторону усиления ихвзаимной зависимости . Этот комплексвключилв себя двигательныеорганеллы - реснички и жгутики . Заэтой трансформациейпоследоваломорфогенетическоепреобразование , которое состояло вразвитии выраженных клеточных асимметрий .С интеграцией взаимозависимыхнуклеоцитоплазмы, митохондрий, ундулиподий- завершилось создание полигеномной структуры эукариотической клетки. Первая эукариотическая клетка была гетеротрофной. Она питалась готовыми органическими соединениями. У этих эукариот возникли процессы митоза, а затеми мейоза. И наконец , приобретение эукариотами способности к фотосинтезу , имело место в процессе становления основной эукариотической организации и после его завершения- последнее событие в зтой эпопее. Симбиозы становились более и более интегрированными .В настоящеевремязависимость каждойорганеллы отпродуктов метаболизма других органеллстала настолько полной ,что только современныеметоды анализапозволяютпроследить метаболическиепути исходных партнеров . Эукариоты разнообразны по своему строению ,но метаболически ониостаются единообразными .Метаболическиеухищрения эукариотбылиразработаны различнымибактериями еще дотого времени , когдаониобъединились,превратившись вхозяинаиегоорганеллы .Прокариотыдостигли большогоразнообразияв способахполучения энергии ивтонкостях биосинтеза .Они оставилислед своего существования задолгодо того , как наоснове эукариотической клетки сформировалисьтакиекрупные организмы ,как животные , обладающиескелетом .
Схема происхождения эукариотических клеток путем симбиоза
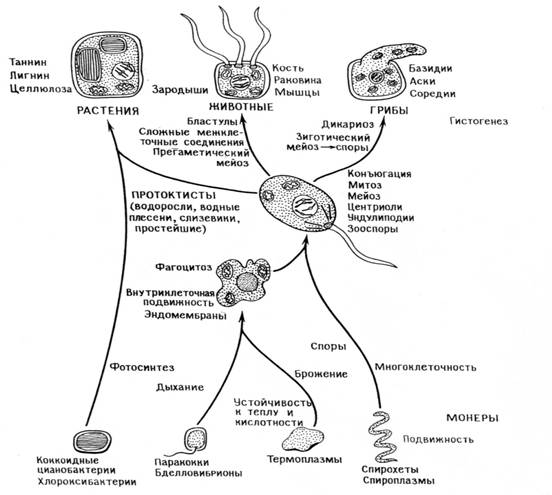
ПРЕДШЕСТВЕННИКИ ЖИЗНИ
Предки нынешнихорганизмоввозникли болеетрех милли-ардовлет назадизорганических соединений ,образовавшихсяна поверхностиЗемли . Привлекательность этой концепции отчасти связана сее доступностью для проверки .
С.МиллериХ..Юри показали ,чтопри атмосферных усло-виях , имевших место на юной Земле ,могут образовыватьсяаминокислоты идругие органические молекулы. Различные типы органическихмолекул образовывались спонтанно в течение первого миллиарда лет существованияЗемли.Многие органические молекулы былиобнаруженыв межзвездном пространствеивметеоритах .Значит , органическиесоединения возникают привзаимодействии легкихэлементов, в при-сутствии источников энергии . Универсальность химического механизма биорепродукции означает , чтовсеживое наЗемлепроисходит отобщихпредков :всянынешняя жизньпроизошлаот клеток синформационнымисистемами , осно-ваннымина репликации ДНК , ина синтезебелков , направляемоминформационной РНК .
ФЕРМЕНТИРУЮЩИЕ ОРГАНИЗМЫ
Возможно ,что нуклеиновые кислотыс самого началабыли отделеныот окружающейсреды липопротеидными мембранами . Информация стала кодироваться последовательностью нуклеотидов вмолекулах ДНК .Древнейшие клеткииспользовали АТФ иаминокислоты ,входящие в состав белков , как прямые источникиэнергии , и какструктурные компоненты . Итак - древнейшая жизнь существовала в формегетеротрофных бактерий , получавшихпищуи энергию изорганического материала абиотического происхождения . Для неодарвиновскойорганической эволюции необходимотрифеномена: воспроизведение, мутации и селективное давлениесреды. Воспроизведение - этонеприменное условиебиологи-ческой эволюции . При достаточно точной репродукции - мутациииестественный отбор неизбежны . Объяснить проис-хождение такой репродукции -значитпонять происхождениесамойжизни .Мутирование- первичныйисточник изменчивостив эволюции , ведеткнаследуемым изменениям в самовоспроизводяйщейся системе .Как только выработалось надежное воспроизведение , началась неодарвиновскаяэволю-ция .Организмы , укоторых выработались пути синтеза нужных клетке компонентов,имели преимуществоперед другими .Древнейшимиавтотрофами ,тоесть организмами способнымиудовлетворять своипотребностив энергии иорганических веществах засчет неорганических источников ,были анаэробныеметанообразующиебактерии . Способность кфотосинтезу впервые выработаласьубактерий , чувствительныхккислороду .Поглощаясолнечный свет с помощьюбактериальногохлорофилла , они превращали угле-кислоту атмосферы в органические вещества.Появление ана-эробного фотосинтезанеобратимоизменило поверхностьиатмосферу Земли.
Углекислотаудалялась изатмосферы и превращаласьвор-ганическое веществовсоответствии собщимуравнением фотосинтеза:2H2X+CO2=CH2O+2X+H2O . Фотосинтезирующиебактерии, которые использовали для восстановления СО2 сероводород, откладывалиэлементарную серу . Онидали началобактериям ,способным к фотосинтезу свыделением кислорода. Вероятноэтот типфотосинтеза зародился врезультатенакопления мутаций у фотосинтезирующих серных бактерий .Этот путьпривелк возникновению сине-зеленыхводорослей илицианобактерий .Недавно былаоткрыта и изучена новая группа фотосинтезирующихпрокариот ,выделяющихО2 - хлороксибактерии . Можнопредполагать ,чтоименно циано-и хлорооксибактерии были предкамипластид,находящихся вклетках водорослей ирастений .Помере накопленияО2 в результате микробного фотосинтеза земная атмосфера становилась окислительной,что вызвалокризис всей жизни – доэтоговремени мир былзаселен анаэробнымибактериями. Чтобывыжитьмикробы должныбылиизбегать кислорода, укрываясьвболотах , илидолжныбыли выработатьобменвеществ , способныйпредохранить ихотвозрастающего количествакислорода .
ФОТОСИНТЕЗ И ВОЗДУХ
Примерно 2 миллиардалетназад одинзадругим происходилиразличныеэволюционные процессыпо созданиюметаболи-ческогоразнообразия . Способностьквыработке специфическихферментовможет передаватьсявпределах одноговидас помощьювирусоподобныхчастиц,называемыхплазмидами. Поразительныбиосинтетическиеспособности прокариот: изуглекислоты , несколькихсолейи восстановленных неорга-ническихсоединений, служащихисточниками энергии , микро-организмы- хемоавтотрофы могутсинтезироватьв содержащейкислороднеосвещенной водевсесложнейшие макромолекулы, необходимые дляростаи размножения . Этибиохимические про-цессыприводилик системамкруговоротаэлементов ещетогда, когдане былорастений, грибов , животных .
Ассоциации и эукариоты .
Помере накопленияатмосферногокислорода абиотическаяпродукцияорганических веществсокращалась , так какони быстроразрушалисьпод действиемэтогогаза .Бактерииоказались впрямойзависимости друготдруга о том , чтокасается притокагазов , вентиляции иудаленияотходов мета-болизмавгазообразном виде .Создаласьих взаимнаязависи-мостьв отношении питанияи защиты ,возникло многоразногорода связеймеждуорганизмами , включаясимбиоз , паразитизм ихищничество. Определеннаясериясимбиозов велакформи-рованию новыхтиповклеток . Согласнотеории последова-тельных симбиозов, всеэукариоты сформировалисьврезуль-тате симбиозамеждучрезвычайно далекимидругот другавидамипрокариот: нуклеоцитоплазма образоваласьиз микро- организмовхозяев , митохондрии избактерий ,дышащихкислородом , пластиды -из хлороокси-илицианобактерий , аундолиподии -из спирохет , прикреплявшихсякповерхности хозяев . Митотическоеделениеклетки выработалосьтолькопосле того, как хозяевастали поглощатьспирохети передислоци-роватьихчасти .Итак , вначалеистории эукариотнаходилисьлишенные клеточнойстенкиплейоморфные микроорганизмы ,способныесбраживать глюкозудодвух- и трех углеродных ко-нечных продуктов по метаболическомупутиЭмбдена-Мейер-гофа. Этиорганизмы , которымпредстоялостать нуклео-цитоплазмой ,приобреталиэндосимбионтов . Акаковаприрода эндосимбионтов , дышавшихкислородом? Эти промитохондриибылиаэробными палочковиднымиорганизмами , близкимиксовременным бактериям ,вродеParacoccusdenitrificans.Ассоциация могла начатьсясхищничества, объектом которогослужиликлетки-хозяева . Размножение симбионтоввнутриос-мотически регулируемойклеткивело котбору,направленному противсохраненияими клеточныхстеноки биосинтетическихпутей,дублирующих путихозяина. Естественныйотбор привелкобразованию крист -складчатых мембран современных митохондрий. Итогомэтогоприспособления к симбиозу было образование двугеномных клеток – аэробных , содержащих митохондриииядра ,но неспособных к митозу ,крупных амебоидных микроорганизмов . Следующимшагом кполномуэукариотическомустатусу было симбиотическое приобретение подвижности .Ундулиподии образовались толькопослетого ,какспирохеты объединились самебоидами инавсегда видоизменились .Как ивовсех устойчивыхассоциациях ,репродукция симбионтовстала протекать согласованно срепродукцией хозяев ипоявились новыекомплексы , содер-жащие тригенома :ядерный,митохондриальныйи ундолипо-диальный .Этитригеномные комплексыдалимножество различныхвидовпротистов . Симбиотические бактериистав-шие ундолиподиями ,обеспечилипреадаптацию кмитозу . Развитие механизмовмитоза привело врезультатеадаптивной радиации кобразованию огромногомножествавидов эукарио-тическихмикроорганизмов .Итак , гетеротрофныемикробы – эукариоты породили животных игрибы . Другая сериясимбиотическихприобретений привела кэукариотам - фотосинтетикам . Естественныйотбор оптимизировал процес-сы митоза, мейоза .Протопластидыначали ассоциироваться с гетеротрофными микрооорганизмами ,которые заглатывали микробов – фотосинтетиков , нонепереваривали их. Прокариоты хлороксибактерии- прародителипластид простей-ших , зеленыхводорослей . Цианобактерии- стали предками красноватых пластид красныхводорослейи криптомонат. Великие новшества в эволюции клеток возниклиещедотого , какпоявилисьпервые животныерастенияи грибы. Какие – то гетеротрофные эукариоты в результатепоглощения фотосинтезирующихпрокариот сталиводорослями . Это завершающеесобытие произошло более 400 млн.лет назад ,очем свидетельствуютордовикские ископаемые остатки крупных фотосинтетиков- сифоновые зеленыеводоросли икрасные водоросли .
Взгляднаэволюцию клетки.
Прямая филиация.
Традиционныйвзгляд напроисхождениеи эволюциюэукарио-тическихклеток заключаетсявтом , что всеживыеорганизмы произошлипрямымпутем отединственнойпредковой популя-цииврезультате накоплениямутацийпод действиеместест-венногоотбора.
Точечныемутации, дупликации,делециии другиенаследствен-ныеизменения , какизвестно игравшиерольв эволюцииживот-ныхи растений , ответственныиза дифференцировку эукарио-тических клетокиз прокариотических-теорияпрямой филиации . т. е. представление онесимбиотическомпроисхождении клеток. Возможно , зеленыеводорослиявляются потомкамиродоначаль-никоввсех растений.Многиеособенности этихводорослейнаводят намыысль , чтоониирастения имеютобщегопредка. Филогения животныхсложнее , но концепцияпрямойфилиации вероятноприложиматакже и кихпутям эволюции . Губкиочевиднопроизошли отпротистов ,независимо отостальныхгрупп животных .Итак , покрайней мередляживотных ирастений гипотеза , покоторой сложныеэукариотические орга-низмы развилисьизболее простоустроенныхэукариот путемнакопленияотдельных мутацийподдействием естественногоотбора , вполнеправомерна .Эволюционные новшества опирают-сяна сохранениевысокосовершенныхблагоприятных генов .Итак , новыеорганизмы сповышеннойспособностью контроли-роватьсвоюсреду обитанияразвивалисьбольше благодарясохранениюдупликации ирекомбинациивысокоадаптированных генов ,чем путемприобретенияновых свойствврезультате простых , случайныхточечныхмутаций . Гипотезапрямойфилиации непозволяетобъяснить происхождениеэукариот от прокариот . Напримерхиатус междунеспособнымик митозуцианобактериями , итакими формамисвполне развитыммитозом , каккрасные водоросли , труднопонятьс позицийпрямойфилиации , но онестественновытекает изсимбиотической теории .
Согласнотеории прямой филиации и теориисимбиоза , все организмы наЗемле произошли от биохимически одно-типных бактериальных предков , арастениям иживотным далиначало эукариотические микробы . Фотосинтезирующие бактерии далиначало водорослям , и вконце концов расте-ниям , а некоторые водоросли утратилисвоипластиды ипре-вратилисьв предков грибов и животных . Теоретикипрямой филиациисогласны втом ,что пластидыидругие органеллыэукариот , включая ядро ,сформировалисьпутем дифферен-циации внутрисамих клетоки называют этоботаническим мифом .
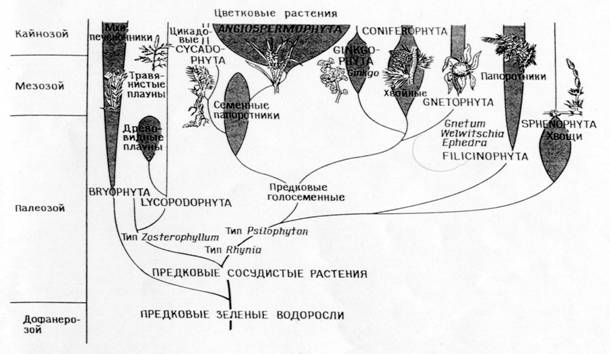
Филогения типов растений
Сопоставление филогений , соответствующих теории прямой филиации (А)
и теории симбиоза (Б)
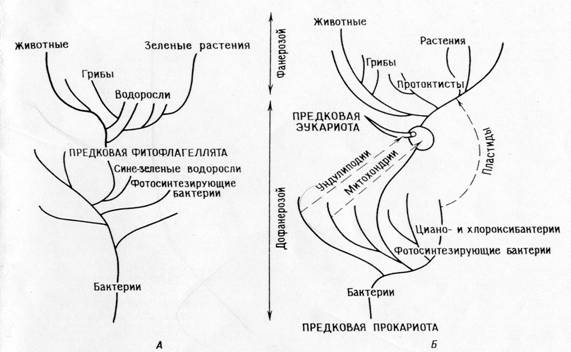
Различия междуположениямитеории прямойфилиациии симбиотической теории:
Теорияпрямой филиации .
Главнаядихотомия:животные – растения
Фотосинтезирующиеэукариоты (водорослии зеленые растения )произошлиот фотосинтезирующихпрокариот ( сине-зеленыхводорослей)
Растенияи ихфотосинтезируюшшиесистемы вдокембрииэволюционировалимонофилетически .
Животныеи грибыпроизошлиот автотрофов , утративщихпластиды .
Митохондриидифференцировалисьвнутриклеточноу фотосинтезирующихправодорослей .
У праводорослейвнутриклеточнодифференцировалисьундолиподии , митотическийаппарат ивсепрочие эукариотическиеорганеллы .
Все организмыпроизошлиот предков – прокариотпутемнакопления одиночныхмутаций .
СЛЕДСТВИЕ:согласнотеории , должнысуществоватьправодоросли; трудносвязатьэту теориюсископаемой летописьюипостроить соответствующуюфилогению .
Теорияпоследовательныхэндосимбиозов:
Главнаядихотомия: прокариоты – эукариоты .
Фотосинтезирующиеэукариоты (ядросодержащие водоросли ирастения ) инефотосинтезирующиеэукариоты ( животные , грибы , простейшие) произошлиот общих гетеротрофныхпредков ( амебофлагеллят ).
Эволюция фотосинтеза вархеебыла монофилетической убактерий ( включаяциано- ихлорооксибактерий ), фотосин-тезирующиепротисты возникливнезапно ,когдаихгетеро-трофныепротисты – предкивступили всимбиозс фото-синтезирующимипрокариотами (протопластидами ).
Животныеи большинствоводорослейи грибовпроизошлинепосредственноот гетеротрофныхпротистов .
Митохондрииуже былиугетеротрофных протистов ,когдапоследниев результатесимбиозаприобрели пластиды .Промитохондриибыли ранееприобретенысимбиотическимпутем .
Митозвыработался угетеротрофныхэукариотическихпротистов путемдифференциацииундолиподиальнойсистемы .
Прокариотыпроизошли отранних прокариотическихпредков путемряда одиночных мутационныхсобытий , эукариотытожепроизошли отэукариотическихпредков благодаряодиночныммутационным событиям , эукариотические же предкипроизошлиот прокариотврезультате сериисимбиозов .
Следствия:теорияобъясняет биохимическиепутии ихизбыточностьу органелл . Онапозволяетпостроить последо-вательнуюфилогению, объясняющую “биологическийразрыв “ междувременами протерозояифанерозоя . Теорияпредска-зывает , чтоу органеллдолжнысохраняться остаткисистемрепликации .
Ботанический миф
Со временЖюсьенизшие растения называютталлофитами .авысшие - бриофитамиитрахеофитами . Согласноботаническому мифу ,общиепредкифотосинтезирующихбактерий исине-зеленыхводорослей путемпрямойфилиации далиначалоэукариотическимводорослям втомчисле морским .Группыорганизмов , филогениюкоторых внаибольшшеймере затра-гиваетконцепцияпрямой филиации, оказалисьв областиботаники -это бактерии , водоросли , грибы ирастения .
Гипотетическиеорганизмы , которые моглибы бытьсвязующимзвеном междуцианобактериямии зеленымиводорослями , былиназваны родоначальнымифито-монадами ( ancestral phytomonads ) или праводорослями .Некоторыеавторы утверждали , чтоподходящимикандитами нарольправодорослей могутбытьэукариоты Cyanidiumи Cyanophora,сходныепо своим пигментнымсистемамс цианобактериями . У таллофитовнебыло обнаруженопромежуточныхформ междунемитотическимсостоянием цианобактерий , ивполнеразвитым типичныммитозомзеленых водорослейирастений . Хотягрибы ицветковыерастения полностьюлишеныундулиподий , имсвойственны митозимейоз , длякоторыхнеобходимы веретена , состоящиеизмикротрубочек .Имеется общеесогласиевтом ,чтовсе многоклеточныеорганизмы ,обладающиемикротрубочками ,происходятот эукариотическихмикроорганизмовс ундолиподиями ,ноникто незнает , откакой именногруппыпротистов . Подкреплениемботаническогомифа служилидревностьи сложностьфотосинтеза , атокже тотфакт , чтоу многихорганизмов ( убактерий , евгленовых , пара-зититических растений )какв природе , такив лаборатории , наблюдалась утратафотосинтезав результатемутаций .Поскольку фотосинтез - анаэробныйпроцесс ,обеспечивающийвконечном итогесуществованиевсех организмов ,тоондолженбыл развитьсяоченьрановистории жизнинанашей планете , ещедотого каквозникмитоз , длякоторого небходим кис-лород .вфотосинтезе участвуетмножествопигментов , липидови ферментов ,упорядоченно расположенныхвочень сложныхмембранах .При этом деталифотосинтетическихмеханизмов уводорослейи растений, содной стороны, иу цианобактерий , сдругой- поразительносходны .Все этиорганизмыосвобождают газообразныйкислородизводы , котораяслужитисточником атомовводорода , используемыхдлявосстановленияСО2 доорганическихвеществ клетки . Многиеводорослии простейщие неимеют нимитоза , ни генетическойсистемы , сходнойстаковой животныхирастений . Инфузорииимеютособую гене-тическуюсистемус ядрами2типов , причемклетки нередкомногоядерные . Одинтип яядер – макронуклеусы , а другой – намногоменьшие микронуклеусы . Микронуклеусы физиологи-ческинесущественны . однако онихранят копиигенов, апри половомпроцессе -претерпевают мейоз .Физиологическинеобходимые макронуклеусыслужатместом транскрипцииРНК, и , такимобразом , управляют физиологиейклетки .Одна изтрудностейпри выясненииотношениймежду цианобактериямииэукариотическимиводорослями связанасотсутствием данныхопроисхождении ундолиподий: небыло найденоникакихпромежуточных звеньевмеждуцианобактериями, у которыхни-когдане бываетундолиподий , имножеством водорослей , которыеихимеют . Другаяпроблема – полноеотсутствие половогопроцессау цианобактерий:между бесполымицианобактериямии водорослями , большинству которыхсвойствененполовой процесс , неоказалосьникаких промежуточныхформ .Обнаружилось , чтобольшинство водо-рослейимеютундолиподии накакой – либостадии жизненногоцикла .Подвижные структурыуводорослей , простейших , жи-вотных ирастений оказалисьодинаковыми: всеони состоятизмикротрубочек диаметром0,024 мкм . Выяснилось ,чтомито-тическоеверетено дажеугрибов , у которыхнетундолипдий ,состоитиз белковыхмикротрубочектого жедиаметра , чтои вундолиподиях .На протяженииполувекаказалось чрезвычайно правдоподобным, чтоэукариотичкскиеводоросли произошлиотцианобактерий . Этотботанический мифпривелк предположе-ниям , которые многиегодыслужили какстимуломдля исследования , такисистемой координат ,вкоторойпро-изводиласьоценка получаемыхрезультатов . Утверждение , чтосвязующиезвенья междуцианобактериямии эукариотическимиводорослямивымерли , не оставивископаемых следовиживых реликтов , становилосьвсеменее убедительнымпомере поступ-ленияновыхданных оживыхмикробах иобокаменелостях . Главныепромежуточные звеньямеждуфотосинтезирующимипрокариотами иэукариотамине вымерлибесследно , кактого требуетботаническиймиф ,- они никогданесуществовали . Согласнотеории последовательныхэндосимбиозов , циано-бактерии – действительнородоначальники ,нотолькопластид , а неостальныхчастей эукариотическойклетки , иуж вовсякомслучае неееядра . И всежев вопросеопроисхождении ядрагипотезапрямой филиацииитеория симбиозасовместимы . Мыслимыемодификации крайнеговарианта теориисимбиозавключают , например ,идеюо симбиотическомпроисхождениипластид , но немитохондрий , илио симбиотическом про-исхождениипластиди митохондрий , нонеундолиподий . Сторонникипрямой филиациивыдвинулитакже предположениеопроисхождении ядрапутемсимбиоза .
Решающиедоказательстваэндосимбиотическогопроисхождения хлоропластовимитохондрий .
Наиболеесущественные доказательствапроисхожденияважней-ших органелл эукариотическихклеток - хлоропластовимито-хондрий изпрокариотполучены впоследниегоды приизуче-нииструктуры рибосомальныхРНК . Молекудярныйбиолог
К. Возпоказал ужев80-х годах , чтоструктураРНК изрибосомхлоропластов растенийпочтиидентична структуреРНКиз некоторыхцианобактерий . Втожевремя группаисследователейустановила , чтоРНК рибосом ,полученныхиз митохондрийразныхэукариотов схожасРНК некоторыхбактерий, в частностибактерийродаParacoccus . Набор дыхательныхферментовуказанной бактерииоченьпохож на
“дыхательныйансамбль “ митохондрийживотных .
Эти данные достаточноясно свидетельствуютвпользу большейвероятностиэндосимбиотическогопроисхождения эукариотическихклеток . Явлениеэндосимбиоза ипосей деньчрезвычайношироко распространеновприроде . Известно , чтонателе некоторыхморскихрыб частопоселяютсясветящиеся бактерии , которыенетолько самиполучаютбольшие преимуществадлясвоей жизни , но иоченьполезны дляорганизмовхозяев . Все сказанноеубедительносвидетельствуетв пользу эндосимбиотическогопроисхождения эукариотическихклетокиз гораздоменьщихпо объемуклетокпрокариот . Однакоглавный вопросзаключаетсяв установленииприродыхозяйской клетки ,ведьклетки эукариотобладаютхорощо сформированнымядром , имеющимоболочку ,а прокариотытакогоядра неимеют . Л. Маргелис , например , считает ,что рольхозяйскихклеток , возможно, сыграли прокариоты – предшественники современныхбактерий , относящиесякмикоплазмам - оченьпримитивнопостроенным прокариотам , практически лишеннымклеточныхстенок . В последнеевремяпоявились иширокообсуждаются работыяпонскогоученого
Т. Ошимы . Наосновании тщательногоизученияструктуры рибосомныхРНКмногих про-иэукариот , в томчислеи многихпредставителейдо сихпормалоизученногонадцарства архей , этотавторприходит квыводу , чтохозяйской клеткойявлялсяодин изпредковсовременных архей . Следуетсказать , чтоу представителейименноэтого надцарства , в отличиеотбактерий , имеетсяоченьмного общихсэукариотами биохи-мическихимолекулярно – биологических особенностей . В по-следнихработах Т. Ошимаприводятсядоводы впользутого , что хозяйскойклеткойпривозникновенииэукариот являлсяпредокархей , подобныйодному изсовременныхпредставителейэтих прокариот-термоплазме . Каковобудет окончательноерешениепроблемы установленияприродыхозяйской клетки , давшейначалоэукариотическойклетке впроцессемножествен-ногоэндосимбиоза , покажутдальнейшие исследования .
ВЫВОДЫ:
Наиболее популярнойиобоснованной гипотезойпроисхожденияэукариотическихклеток являетсявнастоящее времяпредставле-
ниеобих образованиипутеммногократно происходившеговпроцессе эволюциивтечении первых1.5 – 2млрд летсуществованияна Землеживыхклеток эндосимбиозаразличныхпрокариот , относящихсякбактериям иархеям .Решающие доказательства эндосимбиотического происхожденияхлоро-пластови митохондрийбылиполучены ведущимибиологамимира приизученииструктуры рибосомальныхРНК. Этиданныедостаточно ясносвидетельствуютв пользубольшойвероятности эндосимбиотического происхождения эукарио-тическихклетокиз гораздоменьшихпо объему(нанесколько порядков ) клетокпрокариот .
ЛИТЕРАТУРА
1.Соросовский образовательный журнал.Биология , Химия , Науки о земле , Физика , Математика . N 51998г
2 Маргелис Л.Роль симбиозав эволюции клетки. Пер.с англ.,М. МИР1983г
3 Основы микробиологии .М . “Медицина “ 1992 .
4 Хахина Л.Н. КонцепцияА.С. Фаминцынао значениисимбиозав эволюции Л.; Наука , 1981 , с . 165 – 181 .
5 Альбертс Б . Молекулярнаябиологияклетки;Пер. сангл . М . Мир1986 т.1