Векторные системы для молекулярного клонирования в Bacillus subtilis
| Загрузить архив: | |
| Файл: ref-22929.zip (882kb [zip], Скачиваний: 192) скачать |
БЕЛОРУССКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
Биологический факультет
кафедра генетики
ВЕКТОРНЫЕ СИСТЕМЫ ДЛЯ МОЛЕКУЛЯРНОГО
КЛОНИРОВАНИЯ В Bacillussubtilis
Курсовая работа
студента 3 курса КОВАЛЬЧУКА К.В.
Научный руководитель:
канд. биол. наук,
доцент ТИТОК М. А.
Минск 2004г.
ОГЛАВЛЕНИЕ
|
ВВЕДЕНИЕ……………………………………………………………………3
ОБЗОР ЛИТЕРАТУРЫ………………………………………………………..4
1.1 Клонирующие векторы…………………………………………………..4
1.2 Векторы экспрессии……………………………………………………...6
1.3. Векторы для клонирования промоторов………………………………10
1.4 Векторы с регулируемой копийностью………………………………..10
1.5 Векторы для направленной инактивации генов………………………10
1.6 Векторы для получения гибридных белков…………………………...10
1.7 Геномные векторы……………………………………………………....10
ЗАКЛЮЧЕНИЕ………………………………………………………………..10
ЛИТЕРАТУРА………………………………………………………………..10
ВВЕДЕНИЕ
Генетическая инженерия является одним из важнейших направлений биотехнологии, науки об использовании живых организмов и биологических процессов в производстве. По сути, генетическая инженерия (её называют также молекулярным клонированием или технологией рекомбинантных ДНК) представляет собой совокупность экспериментальных процедур, позволяющих осуществлять перенос генетического материала из одного организма в другой. В основе этого переноса, лежитвстраивание нужного фрагмента ДНК (вставки) в другую молекулу ДНК (вектор), которая способна реплицироваться в соответствующей клетке-хозяине.
Исторически сложилось так, что наиболее изученными в генетическом плане являются грамотрицательные бактерии, в частности Escherichiacoli. И соответственно большинство векторных систем разработано именно для этого организма. Однако существует целый ряд бактерий обладающих свойствами, которые не присущи E.coli. Как правило, это грамположительные микроорганизмы, и чтобы изучать и использовать на практике их свойства разрабатываются векторные системы, учитывающие специфику данных микроорганизмов. Одним из таких микроорганизмов являются грамположительные бактерии Bacillussubtilis.
По изученности генетического аппарата клетки Bacillussubtilis находятся на втором месте после E.coli. Для них характерен нечасто встречающийся среди бактерий процесс споруляции,и в этом плане они представляют значительный теоретический и практический интерес. B.subtilis имеют важное практическое значение: они широко используются в различных процессах ферментации при промышленном получении антибиотиков и ферментов и в этом отношении обладают рядом ценных свойств, таких как способность секретировать синтезируемый белковый продукт в культуральную жидкость, способность расти на дешёвых субстратах и обладать продуктивностью на один-два порядка выше, чем у грамотрицательных бактерий-продуцентов.
Поскольку B.subtilisобладают целым рядом описанных выше свойств, для этого организма было разработано множество разнообразных векторных систем. В данной работе расмотрены основные типы векторных систем, с помощью которых можно производить различные генетические манипуляции в клетках B.subtilis.
ОБЗОР ЛИТЕРАТУРЫ
Первоначально большая часть векторов для B.subtilis создавалась на основе геномов умеренных фагов (таких, как rho11 и phi105) , т.к. они обладали большей стабильностью, чем векторы на основе плазмид, реплицирующихся по типу катящегося кольца (rolling-circle-replication, RCR-плазмиды) (Негрук, 1991). В настоящее время большинство векторов конструируется на основе репликонов тета-плазмид, обладающих высокой структурной и сегрегационной стабильностью. Кроме того, в последнее время разработан ряд так называемых геномных векторов предоставляющих дополнительные возможности по сравнению с плазмидными векторами.
1.1. Клонирующие векторы
В штаммах B.subtilisсодержится незначительное число естественных плазмид, причём большинство из них являются криптическими. Однако из других видов рода Bacillus, а также других родов грамположительных бактерий, было выделено множество плазмид, способных к автономной репликации в клетках B.subtilis. Первоначально плазмидные векторы для B.subtilis конструировались на основе мелких (размером меньше 10 kb) плазмид различных грамположительных бактерий, в первую очередь на основе плазмид, выделенных из Staphylococcusaureus(pC194, pE194, pUB110 и др.). Однако векторные молекулы, полученные на основе таких плазмид, проявляли высокую структурную нестабильность, и с их помощью можно было клонировать только короткие сегменты ДНК, а длинные сегменты часто подвергались перестройкам (Michel et al., 1980; Ehrlichetal., 1986).
Нестабильность мелких плазмид в клетках B.subtilis обусловлена механизмом их репликации. Указанные плазмиды реплицируются по типу катящегося кольца (RCR плазмиды), а вследствие разобщённости синтеза лидирующей и отстающей цепей, в процессе репликации образуется одноцепочечная ДНК, которая сильно стимулирует рекомбинацию между гомологичными последовательностями (Ehrlih etal, 1986). Одноцепочечный надрез, образуемый во время инициации репликации, также может быть причиной рекомбинации (Michel, Ehrlich, 1986; Baliesteretal., 1989; Grosetal., 1989). Кроме того, вставки ДНК могут вызвать образование высокомолекулярных конкатемеров, увеличивая число копий плазмиды (Gruss, Ehrlich, 1989; Dabertetal., 1992), что может повысить частоту рекомбинации, а также поспособствовать отбору плазмид, утративших вставки.
Существует также другой тип плазмид, реплицирующихся в соответствии с механизмом тета-типа. При репликации по этому механизму не образуется интермедиатов в виде одноцепочечной ДНК, и по этой причине векторы, созданные на основе тета-репликонов гораздо стабильнее и, следовательно, эффективнее векторов на основе RCR плазмид. Первые данные об этом были получены в экспериментах по получению и изучению свойств клонирующих векторов pHV1431, pHV1432, pHV1435 и pHV1436 (Janniereetal., 1990).
Челночный вектор pHV1436 (рис. 1) был создан на основе репликона плазмиды pTB19. На основе репликона pAMb1 были сконструированы три клонирующих вектора: pHV1431, и полученные из него pHV1435 (путём инверсии rep-области pAMb1)и вектор pHV1432 (путём делеции небольшого фрагмента в последовательности rep-области pAMb1).

Рис. 1. Структура клонирующих
векторов. oripTB52
и oripAMb1,репликационные регионы природных плазмид pTB19 и pAMb1
соответственно; pBR322,
последовательность pBR322,
включающая её репликационный регион; 
Данные векторы оказались структурно намного более стабильными, чем векторы на основе сигма-репликонов (Janniereetal., 1990):
(1) при их использовании наблюдается гораздо более низкая частота рекомбинации между повторяющимися последовательностями (снижена в 1000 раз)
(2) с их помощью можно клонировать значительно более длинные сегменты ДНК; средний размер вставки 10 и 1 kb, с диапазоном 3 -17 kb и0.1 - 1.7 kb для векторов на основе тета и сигма репликонов соответственно
(3) в отличие от RCR плазмид они способны к стабильному поддержанию больших сегментов ДНК
Данные свойства характерны и для других векторов, являющихся производными тета-плазмид. Кроме того, было показано, что в отличие от сигма-плазмид, тета-репликоны, со вставками или без них, не образуют или образуют незначительное число высокомолекулярных конкатемеров (Gruss, Ehrlich, 1988; Dabertetal., 1992a; Dabertetal., 1992b). Это также является одной из причин их структурной стабильности.
Помимо клонирующих векторов для Bacillussubtilis был создан ряд векторов специального назначения: векторы экспрессии, клонирования промоторов и терминаторов, векторы с регулируемой копийностью и т.д. При этом используются векторы на основе как сигма- так и тета-репликонов.
1.2. Векторы экспрессии
В простейшем случае для экспрессии достаточно клонировать нужный ген вместе с его промотором и сайтом связывания с рибосомой (ribosomebindingsite, RBS) в природную плазмиду, способную реплицироваться в хозяине, в котором нужно проэкспрессировать данный ген. Так, например, в B.subtilis был проэкспрессировать ген термостабильной арабиназы термофильной бактерииBacillusthermodenitrificans. Для этого фрагмент, содержащий данный ген (с предполагаемым промотором и RBS) был клонирован в природную плазмиду pUB110 (Takao etal., 2002).
Иногда удобно экспрессировать ген с не своего промотора, что позволяет вести экспрессию на более высоком уровне, или контролировать её. Так, ген термоактивной пуллуланазы гипертермофильной анаэробной архебактерии Desulfurococcus mucosus (apuA) был успешно экспрессирован в B.subtilis(Duffneretal., 2000). Экспрессия гена шла под контролем промотора PamyM (рис. 2).

Рис. 2. Структура вектора pJA803 для клонирования пуллуланазы. RepB - область репликации pUB110; Cm - ген устойчивости к хлорамфениколу; apuA - ген пуллуланазы; PamyM - промотор гена мальтогенной a-амилазы из B. stearothermophilus.
В обоих вышеописанных случаях наблюдался относительно невысокий уровень экспрессии, однако, белки секретировались в среду в количестве, позволяющем их выделение в чистом виде и дальнейшее изучение свойств.
В случае, когда необходимо регулировать уровень экспрессии или экспрессировать белок в очень больших количествах, используют более сложные системы экспрессии.
B.subtilis является очень удобным организмом для проведения в нём экспрессии различных продуктов, т.к. эта бактерия обладает целым рядом полезных свойств: непатогенность, наличие механизмов секреции, хорошо изученные генетика и условия необходимые для экспрессии генов, простотаманипуляций с использованием стандартных протоколов. Для повышения уровня продукции гетерологичных белков в клетках B.subtilis используют штаммы, дефектные по протеазам. Кроме того, можно применить более эффективные регуляторные элементы транскрипции и трансляции. Так, например, была создана кассета Veg(Lam etal., 1998) содержащая сильные регуляторные элементы, подходящие для эффективной экспрессии и секреции гетерологичного белка. В состав кассеты вошли: промотор B.subtilis vegI;lac оператор E.coli; сайт связывания с рибосомой(RBS) B.subtilis; лидерная последовательность стафилококкового белка А (SPA); терминатор транскрипции глюконатового оперона B.subtilis(gnt); полилинкер(multiplecloningsite, MCS), содержащий сайты рестрикции XbaI, SmaI; стоп кодоны для всех рамок считывания; фланкирующие EcoRI и KpnI сайты (рис.3).

Рис. 3. Схематичное изображение регуляторных элементов в кассете Veg.
Длина кассеты356 п.н. RBS - сайт связывания рибосомы; SPA - стафилококковый белок А; MCS - полилинкер. Подробности в тексте.
Кассета Veg была вставлена в полилинкер челночного вектора pM2 (рис. 4) с образованием pM2Veg – вектора для экспрессии и секреции белка в B.subtilis.
Для проверки эффективности функционирования вектора pM2Veg , в качестве гена-репортёра применили ген эндоглюконазы (ген cenA из Cellulomonasfimi). Была осуществлена вставка данного гена в сайт XbaI вектора pM2Veg. С помощью данной системы была достигнута продукция эндоглюконазы на уровне более чем в четыре раза превышающем самый высокий из уровней продукции достигнутых с помощью других систем экспрессии. Экспрессия и секреция белка оставалась на стабильном уровне, и не оказывала ни каких неблагоприятных эффектов на стабильность вектора и жизнеспособность клеток.
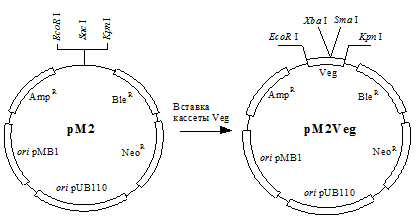
Рис. 4. Схематичное изображение векторов pM2 и pM2Veg.
oripMB1
и oripUB110
- репликационные регионы плазмид pMB1 и pUB110,
соответственно; 

 -гены устойчивости к ампициллину, блеомицину и
неомицину, соответственно.
-гены устойчивости к ампициллину, блеомицину и
неомицину, соответственно.
С помощью pM2Veg в B.subtilis также экспрессирован ген hEGF – ген фактора роста эпидермиса человека. Регистрируемый уровень hEGF был вполне сопоставим с уровнем экспрессии данного гена при использовании других систем. Эти данные указывают на то, что вектор pM2Veg может успешно использоваться для экспрессии в B.subtilisразнообразных гетерологичных белков.
Ещё одна эффективная система экспрессии разработана на основе ксилозного оперона (Bhavsaretal., 2001).
Созданная система экспрессии состоит из следующих элементов (рис.5b):
1) PxylA, под контролем которого идёт экспрессия клонированного гена;
2)¢- концевой участок гена xylA(ген изомеразы ксилозы), содержащий оптимизированный CRE (catabolite-responsiveelement) - цис-действующий элемент, репрессирующий транскрипцию xyl оперона;
3)xylO, с которым в отсутствие индуктора (ксилозы) связывается репрессор XylR;
4)xylR, кодирующий репрессор XylR, со своим промотором PxylR.
Система была вставлена в вектор pDG364 (Cutting, Horn, 1990), способный интегрировать с хромосомой B. subtilis в сайте amyE путём двойного кроссинговера. Таким образом, полученный вектор, обозначенный как pSWEET, позволяет осуществлять встраивание ксилозной системы экспрессии в хромосому B.subtilis. Для определения эффективности функционирования данной системы экспрессии к её проксимальному концу был пришит репортёрный ген bgaB – ген термостабильной b-галактозидазы из B.stearothermophilus (полученная плазмида обозначена как pSWEET-bgaB) (рис.5b). bgaB можно вырезать вместе с RBS с помощью PacIи одной из рестриктаз для полилинкера, и заменить его любым интересующим геном.
|
|
|

|



Рис. 5. a) Структура плазмиды pSWEET-bgaB. AmyE - сайты через которые идёт рекомбинация и интеграция с хромосомой; Cm и Amp - гены резистентности к хлорамфениколу и ампициллину, соответственно; ori - область репликации в E.coli; PacI, сайт рестрикции; b) ксилозная система экспрессии крупным планом. Показаны: ген ксилозного репрессора xylR; промоторы генов xylR(PxylR) и xylA(PxylA ) и оператор xylO;урезанный xylA(первые 58 п.н. + TAA), включающий в себя оптимизированный CRE; сайт связывания рибосомы гена tagD (RBS); ген bgaB, кодирующий термостабильную b-галактозидазу.
Данная система экспрессии
(далее система xyl)
была сравнена с широко применяемой системой экспрессии spac, основанной на применении
лактозных репрессора и оператора E.coli
для контроля экспрессии в B.subtilis. Для этого был
использован вектор pSPAC-bgaB, созданный путём
вставки bgaB в
плазмиду pDR67(Weickert, Chambliss, 1990), содержащую spac систему для
интеграции в сайт amyE.
Для обеих систем экспрессии сайты связывания с рибосомой и окружающие
генетические элементы были одинаковые. Опыты показали, что
1)для системы xyl
характерна значительно более высокая разница между уровнем синтеза
продукта при индукции и при репрессии (экспрессия при индукции в 279 раз выше,
чем при репрессии; для spac
системы – в 16 раз); 2) уровень экспрессии при индукции в 16 раз выше, чем у spac системы; 3) в
отсутствие индуктора уровень экспрессии ниже, чем у spac системы;4)система xyl проявляет чувствительность к
большему диапазону концентраций индуктора.
Была протестирована способность системы xyl обеспечивать ксилоз-зависимую комплементацию. Была проведена попытка комплементировать термочувствительные мутанты (tag-12) по tagD гену, кодирующему фермент, участвующий в синтезе тейхоевых кислот (глицерол-3-фосфат цитидилтрансфераза). Ген tagD дикого типа был клонирован в pSWEET (с образованием pSWEET-tagD) для экспрессии под контролем системы экспрессии xyl. Мутанты с интегрированной в amyE с помощью pSWEET-tagD системой экспрессии и геном tagD под её контролем были способны к росту при непермессивной температуре в присутствии индуктора (ксилозы). Т.е. при росте при непермиссивной температуре происходила ксилоз-зависимая комплементация данного температур зависимого мутанта tag-12.
Таким образом было показано, что pSWEET является эффективной системой для точно регулируемой экспрессии клонированных генов в B.subtilis и, в частности новой эффективной системой экспрессии для условной комплементации в B.subtilis.
1.3. Векторы для клонирования промоторов
Наиболее удобные и универсальные системы созданные для изучения активности промоторов invivo представляют собой плазмиду, несущую ген лишённый промотора,кодирующий продукты, легко поддающиеся количественному анализу (ген-репортёр). Фрагмент ДНК, чью промоторную активность необходимо проанализировать, можно вставить перед геном репортёром. По выходу продукта гена затем можно определить активность данного промотора. Для грамположительных бактерий сконструировано ряд плазмидных векторов для измерения активности промоторов, где в качестве гена-репортёра выступает ген b-галактозидазы или хлорамфеникол ацетилтрансферазы (без промоторов). Главным недостатком этих векторов является их высокая копийность, что мешает их использованию для точного измерения активности промотора invivo. кроме того, они часто имеют узкий круг хозяев.
Создан мобилизуемый малокопийный челночный вектор pTCV-lac(PoyartC., Tieu‑Cuot., 1997) с широким кругом хозяев.
Вектор сконструирован на основе двух репликонов - pACYC184 и pAMb1 (рис.6), что позволяет ему реплицироваться в клетках E.coli и в широком круге грамположительных бактерий (Bacillus, Enterococcus, Listeria, Streptococcus). Точка начала переноса плазмиды RK2 обеспечивает способность кпереносу при конъюгации из клетки-донора E.coli в различные грамположительные бактерии. Роль гена-репортёра выполняет ген b‑галактозидазы (без промотора). Вектор присутствует в клетках в количестве 3-5 копий на хромосому; данная малокопийность позволяет точно измерять активность промоторов.

Рис. 6. Структура вектора pTCV-lac. oriRpACYC184 и oriRpAMb1 - rep-области плазмид pACYC184 и pAMb1, соответственно; ermB - ген устойчивости к эритромицину; aphA-3 - ген устойчивости к канамицину; lacZ безпромоторный ген, кодирующий b-галактозидазу, с сайтом связывания рибосомы для грамположительных бактерий (spoVG); oriTRK2, точка начала переноса плазмиды RK2.
Cконструированный вектор был использован для сравнения активности четырёх промоторов(Ptac, Ptrc, Pspac, PaphA-3) в двух грамположительных палочках (B.subtilis и Listeriamonocytogenes) и двух грамположительных кокках (Enterococcusfaecalis и Streptococcusagalactiae). Для этого осуществлялась вставка фрагментов EcoRI-BamHI в линеаризованный путём рестрикции рестриктазами EcoRI и BamHI вектор. Во всех данных хозяевах производные вектора оказались стабильными. Показано, что активность исследуемых промоторов, измеренная путём определения активности b-галактозидазы, сильно варьирует в зависимости от вида хозяина, в котором исследуется активность данного промотора. Эти данные показывают, что вектор pTCV-lac можно успешно использовать для анализа регуляции генов в широком круге грамположительных бактерий.
1.4. Векторы с регулируемой копийностью
Сконструированы векторы для грамположительных бактерий, позволяющие уменьшать количество их копий на клетку (Renault etal., 1996). Эти векторы созданы на основе pILnew - малокопийного вектора, построенного на основе репликона pAMb1, несущего ген резистентности к эритромицину и способного реплицироваться в большинстве грамположительных бактерией. Этот вектор, как и все полученные из него производные, несут репликационный регион pAMb1, содержащий необходимый для репликации ген repE(кодирует белок репликации RepE) и его регулятор copF. Ген copF был инактивирован путём введения вставки в уникальный сайт KpnI. Так как продукт copF оказывает репрессирующее действие на экспрессию repE, его инактивация ведёт к увеличению копий плазмид на клетку примерно в 20 раз. Первоначальное состояние малокопийности может быть восстановлено с помощью удаления вставки путём его вырезания из KpnI и последующей сшивки. Вектор pILnew был использован для создания (1) клонирующих векторов, (2) векторов для изучения регуляции генов путём клонирования их промоторов и сайтов связывания с рибосомой, (3) векторов для экспрессии генов, (4) кассет, содержащих репликон с различными полилинкерами, которые способствуют созданию новых векторов.



Рис. 7. Структура векторов. Обозначения сайтов рестрикции: E (EcoRI); B (BssHII); N (NdeI); K (KpnI); B (BglII). Em и Cm – гены устойчивости к эритромицину и хлорамфениколу, соответственно. repE – ген, кодирующий необходимый для репликации белок RepE; orfD – ген-регулятор транскрипции repE; сopF - ген репрессора (CopF) транскрипции кластера repE‑orfD; PorfD-repE- промотор кластера repE-orfD; Pres - промотор гена резольвазы resb. MCSpBS-часть MCS плазмиды pBluescript; oriC - репликационный регион pBluescript.
Для того чтобы с помощьюpILnew можно было осуществлять клонирование, в него был введён полилинкер в различной ориентации с образованием клонирующих векторов pJIM2278 и pJIM2279.
Сконструирован ряд векторов, позволяющих измерить активность промоторов с помощью бактериального люциферазного гена из Vibrioharvei в качестве гена-репортёра. Данный ген-репортёр обладает рядом полезных свойств: высокая чувствительность, возможность быстрого измерения активности, отсутствие фоновой активности в грамположительных бактериях. Были использованы два варианта генов люциферазы. В первом случае применялся кластер генов luxA и luxB (luxA/B), во втором - продукт слияния генов (luxAB). Преимущество системы luxA/B над luxAB заключается в её высокой термоустойчивости (luxAB инактивируется при температуре выше 37°C) и в отсутствии токсичности в E.coli, что позволяет создавать конструкции в этом удобном промежуточном хозяине перед их переносом в окончательного хозяина. Однако эти гены могут слабо транслироваться в некоторых грамположительных бактериях, и использование слитых luxAB генов значительно улучшает чувствительность системы в некоторых хозяевах.
Векторы pJIM2366 и pJIM2367 содержат luxA/B гены дистальнее полилинкера и двунаправленный терминатор hisоперонаLactococcuslactisдля предотвращения транскрипции с проксимально расположенных промоторов. В данных векторах было успешно протестировано ряд промоторов из L.lactis при низком и высоком числе копий вектора (промоторы оперонов mle, his и ald из L.lactis).
Активность многих генов регулируется на уровне трансляции. Поэтому была создана версия luxAB гена, позволяющая изучать эффективность сайтов связывания с рибосомой (RBS) для инициации трансляции. Для этого на уровне инициаторного кодона гена luxABбыл введён сайт NdeI, что позволило осуществлять вставку RBS сразу перед стартовым кодоном этого гена. Транскрипция гена luxAB при этом может инициироваться с промоторов, расположенных проксимальнее. Вектор pJIM1715, содержащий слитый ген luxAB без RBS не проявлял люциферазную активность. В данный вектор была осуществлена вставка различных фрагментов гена aldB L.lactis, RBS (конструкция pJIM1723), RBS и анти-RBS (pJIM1728), а также RBS и промотор данного гена (pJIM1726). Данные, полученные с помощью указанных конструкций, показали, что pJIM1715 можно успешно использовать для изучения трансляционной эффективности RBS.
Часто необходимо, чтобы синтез продукта гена шёл на определённом уровне. Для достижения этого можно использовать регулируемые промоторы различной силы. Изменение дозы гена предоставляет дополнительный уровень контроля экспрессии гена. Чтобы показать, что для этой цели можно использовать производные pILnew, в pJIM2279 была осуществлена вставка lux генов и была измерена люциферазная активность при низком и при высоком числе копий вектора. При этом сначала измерялась активность экспрессии с промоторов расположенныхв pJIM2279 проксимальнее MCS (промотор кластера orfD-repE и промотор гена резольвазы), а затем с промоторов клонированных в pJIM2366 и pJIM2367(см. выше). Данные показали, что наблюдается чёткая корреляция между дозой гена (количество копий вектора на клетку) и уровнем люциферазной активности. Был сконструирован клонирующий вектор pJIM2375, содержащий промотор mleS для конститутивной экспрессии на среднем уровне в L.lactis, RBS и сайт для клонирования NdeI.
На основе pILnew были созданы производные векторов pJIM2278 и pJIM2279, в которых репликон pILnew может быть отделён от маркера резистентности путёмразреза одной рестриктазой в уникальном сайте рестрикции. На основе этого репликона можно затем создавать новые векторы несущие различные маркеры или гены-репортёры.
pJIM2244 и pJIM2245 являются производными pJIM2279, содержащими гены устойчивости к эритромицину и хлорамфениколу соответственно. В этих конструкциях из всего полилинкера pJIM2279 оставлен единственный сайт EcoRI (в pJIM2245 также часть MCS плазмиды pBluescript). pJIM2241 и pJIM 2246 являются EmR и CmR производными pJIM2278 соответственно и содержат большую часть полилинкера pJIM2278 на конце BssHII фрагмента содержащего репликон. Первые две плазмиды созданы для восстановления репликона pILnew без дополнительных сайтов рестрикции. Другие две плазмиды позволяют менять маркер, в то время как гены, клонированные в полилинкере, остаются связанными с репликоном.
1.5. Векторы для направленной инактивации генов
Функции не охарактеризованных генов можно изучить путём их инактивации и последующего наблюдения эффекта инактивации при различных условиях роста клетки. Для проведения инактивации генов в B.subtilis создан набор векторов для осуществления инсерционного мутагенеза в хромосоме бактерии (Vagneretal.,1998). Данные векторы, обозначенные как pMUTIN, построены на основе репликона ColE1 (рис.8), поэтому они не способны реплицироваться в конечном хозяине (B. subtilis), что необходимо для проведения инсерционного мутагенеза. Они содержат ген-репортёр lacZ для измерения активности промотора исследуемого гена. Также в векторах присутствует индуцибельный промоторPspac, необходимый для контроля экспрессии генов, находящихся в одном опероне с исследуемым геном и расположенным дистальнее от него. Данный промотор необходим из-за возможного полярного эффекта вставки. Промотор Pspacбыл модифицирован для лучшего контроля его активности. Он репрессируется репрессором LacI (кодируется геном lacI, присутствующим в векторе) и, следовательно, может быть индуцирован IPTG, инактивирующим LacI.
|
pMUTIN |
терминатор |
оператор |
MCS |
|
1 |
t 0 |
O1 |
есть |
|
2 |
t 1t 2 t 0 |
O1 |
нет |
|
3 |
t 0 |
oid |
есть |
|
4 |
t 1t 2t 0 |
oid |
нет |
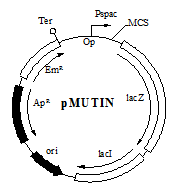
Рис. 8. Структура векторов pMUTIN. Общие черты векторов pMUTIN показаны на рисунке. Различия между векторами pMUTIN указаны в таблице справа от рисунка. Белые боксы являются генами экспрессируемыми в B.subtilis, чёрные боксы - гены экспрессируемые в E.coli. ori - область репликации ColE1. ApR и EmR, гены резистентности к ампициллину и эритромицину соответственно. lacZ, модифицированный ген b-галактозидазы- ген lacZcRBS гена spoVGB.subtilis и стоп кодонами в трёх возможных рамках считывания; Pspac, индуцируемый промотор, содержащий последовательности распознавания РНК-полимеразой фага SPO1 и один из lac операторов - O1 или оператор oid; lacI, ген репрессора LacI модифицированный для конститутивной экспрессии в B.subtilis; Ter - терминаторы: lt0 присутствует во всех векторах ; t1t2 (сильные терминаторы из rrnB) в pMUTIN2 и pMUTIN4. Op - оператор LacI между промоторными последовательностями Pspac: O1 оператор присутствует в pMUTIN1 и pMUTIN2; оператор oid присутствует в pMUTIN3 и pMUTIN4. Различия между векторами pMUTIN указаны в таблице справа от рисунка.
Исследуемый ген можно инактивировать в результате одиночного
кроссинговерамежду вставкой,
осуществленной в вектор pMUTIN
и геном, локализованным на бактериальной хромосоме (рис.9). Для этого
внутренний фрагмент изучаемого гена амплифицируется с помощью ПЦР, клонируется
в полилинкер pMUTIN, и
получившаяся конструкцию используется для трансформации клеток B.subtilis. В результате
интеграции ген прерывается и происходит транскрипционное слияние между его
промотором и lacZ
геном-репортёром. Если данный ген является частью оперона, то остальные
дистально расположенные гены оказываются под контролем промотора Pspac. При этом
терминатор l Pspac предотвращает транскрибирование этих генов с других
проксимально расположенных промоторов. Данные манипуляции приводят к
образованию сразу двух типов мутантов: нулевых мутантов (по orf2) в результате разрыва
гена и условных мутантов (по orf3)
зависимых от IPTG.
Pspac предотвращает транскрибирование этих генов с других
проксимально расположенных промоторов. Данные манипуляции приводят к
образованию сразу двух типов мутантов: нулевых мутантов (по orf2) в результате разрыва
гена и условных мутантов (по orf3)
зависимых от IPTG.

Рис. 9. Интеграция pMUTIN висследуемый ген. Гены оперона orf1-orf3 показаны в виде белых боксов. Заштрихованный ген соответствует внутреннему сегменту исследуемого гена. Вектор интегрируется в orf2 путём одиночного кроссинговера. Стрелки обозначают направление транскрипции, изломанные стрелки обозначают промотор оперона и Pspac.
Созданы векторы для проведения инсерционного мутагенеза в штаммах вида Bacilluscereus, основанного на встраивании мобильного элемента IS231A в случайные сайты хромосомы бактерии (Leonardetal.,1998). Созданные векторы pGIC055 иpGIC057 представляют собой версии одной плазмиды и построены на основе термочувствительного репликона pHT1030. В своём составе они имеют:
1) mini-IS231A, включающий в себяселективный маркер (устойчивость к канамицину [pGIC057] или спектиномицину [pGIC055]) и полилинкерымежду инвертированными повторами;
2)ген транспозазы IS231A (TnA), находящийся под контролем сильного промотора Pdeg;
3) дополнительный маркер- ген устойчивости к эритромицину.
Транспозицию mini-IS231A из плазмид pGIC055 или pGIC057 в хромосому осуществляли при температуре 28°С. Плазмиды затем элиминировали путём повышения температуры культивирования до 46°С. Выяснилось, что в хромосоме Bacilluscereus имеется один сайт сильно предпочтительный для транспозиции mini-IS231A (левый инвертированный повтор транспозона Tn4430). Если в данный сайт уже осуществлена вставка, последующие вставкиосуществляются в случайные сайты.
Путём повторной вставки были получены ряд ауксотрофных мутантов. С использованием рестрикционного анализа данные мутации (также как и предпочтительный для транспозиции сайт) были локализованы на карте хромосомы B.cereus.

Рис. 10. Структура векторов pGIC055 и pGIC057. ori - репликативный регион pHT1030; TnpA-ген транспозазы; MCS 2 - полилинкеры; Pdeg - сильный промотор; инвертированные повторы обозначены черными треугольниками; детерминанты устойчивости: EmR- к эритромицину, mini-IS - к спектиномицину (pGIC055) или канамицину (pGIC057).
Из мутанта ауксотрофного по аденину была вырезана вставка mini-IS231A вместе с прилегающими участками путём рестрикции EcoRI (в mini-IS сайт для данной рестриктазы отсутствует). Путём секвенирования и сравнения с последовательностями из баз данных было выявлено, что сайт, в который была осуществлена вставка, имеет высокую степень гомологии с генами pur оперона B.subtilis, что может указывать на то, что вставка mini-IS231A произошла в один из генов синтеза пуринов. Данные результаты показывают, что рестрикционный анализ вместе с секвенированием позволяют создавать объединённые физические и генетические карты бактериальной хромосомы B.cereus.
Данную систему можно использовать для проведения инсерционного мутагенеза также в других грамположительных бактериях, способных расти при температуре выше 45°.
В геноме B.cereus(как и в большинстве других про- и эукариотических геномов) отсутствует сайт рестрикции I-SceI. Данный сайт присутствует в полилинкере mini-IS. Поэтому последовательное введение двух mini-IS, несущих два разных маркера резистентности, в хромосому приводит к получению фрагмента ограниченного двумя сайтами I-SceI. Таким образом, путём рестрикции I-SceI можно получать фрагменты, подходящие для секвенирования. Данный метод позволяет избежать ряда трудностей встречающихся при использовании традиционных методик клонирования.
1.6. Векторы для получения гибридных белков
B.subtilis долгое время изучается как образец простой клеточной дифференцировки т.к. при голодании у неё запускается программа дифференциальной экспрессии генов называемая споруляцией. Изучение споруляции в свою очередь привели исследованию процессов происходящих в течение клеточного цикла, в частности клеточного деления, сегрегации хромосом и синтеза макромолекул. В проведении этих исследований важную роль играет использование зелёного флуоресцирующего белка GFP (greenfluorescentprotein), применяемого в качестве цитологического маркера.
Слияние кодирующего его гена gfp с другими генами позволяет получать гибридные белки, локализацию которых можно проследить в живых клетках. Для этой цели было разработано ряд векторов (LewisP.J, Marston, 1999; FeuchtA., Lewis, 2001; Kaltwasseretal.,2002).
Была сконструирована серия векторов, несущих разные варианты гена gfp: gfpmutI(pSG1151, pSG1164, pSG1154, pSG1729) и gfpuv(pSG1156, pSG1170) (LewisP.J, Marston, 1999; рис.11a, описание ниже). Продукты данных генов (GFPmut1 и GFPuv) представляют собой мутантные белки, обладающие по сравнению c природным GFP большей яркостью и различающиеся по спектру излучения. Последнее свойство позволяет параллельно проследить локализацию в одной клетке молекул двух типов белков сшитых с разными формами GFP (т.е. проводить двойное мечение клеток).
Векторы pSG1151 и pSG1156 (рис.11a). При использовании этих векторов участок гена, кодирующий С-концевой фрагмент исследуемого белка, вставляется перед gfp. В результате интеграции в хромосому путём одиночного кроссинговера происходит слияние gfp с нужным геном и гибридный белок экспрессируется с промотора данного гена. Проксимальнее вставленного вектора будет находиться неполная копия данного гена. При клонировании целого гена с его промотором в вектор, экспрессироваться будет как гибридный белок, так и белок дикого типа. Это необходимо если исследуемый белок является необходимым для клетки, а гибридный белок не полностью функционален.
Векторы pSG1164 и pSG1170 сходны с описанными выше. Однако они несут индуцируемые промоторы: ксилоз-зависимый Pxyl(pSG1164) и IPTG-зависимый Pspac(pSG1170). Это позволяет контролировать экспрессию дистальнее расположенных хромосомальных генов, если вставка произошла внутрь оперона.
Векторы pSG1154 и pSG1729 созданы для интеграции слитых генов в сайты amyE хромосомы B.subtilis путём двойного кроссинговера. Нужный ген сшивается с последовательностью гена gfp кодирующей N-конец (pSG1154) или C-конец GFP (pSG1729). В обоих случаях для получения функционального гибридного белка необходимо клонировать всю кодирующую последовательность гена. Экспрессия гибридного белка находится под контролем ксилоз-индуцируемого промотора Pxyl.
|



|
FP |
вектор |
|
GFPmut1 |
pSG1151 |
|
GFPuv |
pSG1156 |
|
CFP |
pSG1186 |
|
YFP |
pSG1187 |
|
dsRed |
pSG1194 |
|
FP |
вектор |
|
GFPmut1 |
pSG1129 |
|
CFP |
pSG1190 |
|
YFP |
pSG1191 |
|
FP |
вектор |
|
GFPmut1 |
pSG1154 |
|
CFP |
pSG1192 |
|
YFP |
pSG1193 |


|

Рис.11.a) Векторы, сконструированные для получения белков, слитых с флюоресцирующими белками. MCS - различные полилинкеры; ori - репликативный регион;f1 ori, место начала генерации одноцепочечной ДНК; Cm, Amp, Spc - гены устойчивости к хлорамфениколу, ампициллину и спектиномицину, соответственно. FP - ген флуоресцентного белка (тип флуоресцентного белка указан в таблице под соответствующим вектором; для pSG1164: FP=gfpmut1; pSG1170: FP=gfpuv). lacZ - ген b-галактозидазы;lacI - ген репрессора lacI; amyE - сайты интеграции в хромосому; Pxyl и Pspac - ксилоз-зависимый и IPTG-зависимый промоторы, соответственно; Ppen - промотор пенициллиназы; b) вектор pSG1170, предназначенный для проведения слияния белка клеточного деления DivIVA с GFPuv, после интеграции в хромосому.
Для проверки эффективности работы векторов, вектор pSG1170 был использован для выявления локализации белка DivIVA, участвующего в выборе сайта деления (рис.11b). Как и предполагалось, белок локализовался на обоих полюсах и в центре делящихся клеток.
Было получено ряд производных данных векторов основанных на голубом (CFP, cyanfluorescentprotein) и жёлтом (YFP, yellowfluorescentprotein) спектральных вариантах GFP, а также красном флуоресцирующем белке (DsRed), не являющимся производным GFP(FeuchtA., Lewis., 2001). В первую очередь они были созданы для проведения более эффективного множественного мечения клеток.
Векторы pSG1186 (cfp), pSG1187 (yfp) и pSG1194 (dsRed) (рис.11a) являются производными pSG1151 со следующими изменениями: (i) гены флуоресцентного белка с повышенным содержание GC пар; (ii) изменён полилинкер, расположенный проксимальнее гена флуоресцентного белка; (iii) дополнительный кодон GTG сразу после старт-кодона гена флуоресцентного белка.
Векторы pSG1190 (cfp) и pSG1191 (yfp) являются производными pSG1729 со следующими изменениями: (i) гены флуоресцентного белка с повышенным содержание GC пар (ii) дополнительный кодон GTG сразу после старт-кодона гена флуоресцентного белка. Полилинкеры без изменений.
Векторы pSG1192 (cfp) и pSG1193 (yfp) являются производные pSG1154 с изменённым полилинкером (как у pSG1186 и pSG1186) и дополнительным кодоном GTG сразу после старт-кодона гена флуоресцентного белка.
Данные векторы были использованы для двойного мечения спорулирующих клеток. Была прямо показана последовательная агрегация/дезагрегация белков участвующих в генерации клеточной асимметрии во время споруляции (белки FstZ и SpoIIE). Не смотря на сходность локализации во время споруляции, сигналы от FstZ-CFP и SpoIIE-YPF были чётко различимы друг от друга. В то же время было показано, что природный белок DsRed малопригоден для мечения клеток B.subtilis.
Для характеристики продукта исследуемого генанужны антитела, с помощью которых можно осуществить обнаружение данного белка. Для получения антител необходимо очистить исследуемый белок и иммунизировать им животное. Эта процедура длительная и дорогая, а полученные антитела часто различаются по своим свойствам. Чтобы обойти эти проблемы был создан набор векторов для пришивания к белкам антигенных детерминант (реагирующая с антителом часть антигена) (Kaltwasseretal., 2002): FLAG (искусственно синтезированный пептид), HA (получен из гемагглютинина вируса гриппа) и c-Myc (получен из протоонкогена c-myc человека) длиной 8, 9 и 10 аминокислотных остатков соответственно. Антитела, специфически распознающие эти антигенные детерминанты, имеются в продаже.
a |
|
b |
 |


Рис.12. Векторы для получения гибридных белков. (a) Векторы для получения белков сшитых с антигенными детерминантами. (b) Векторы для получения белков слитых с флюоресцирующими белками. ori - репликативный регион плазмиды ColE1; EmR и ApR, гены резистентности к эритромицину и ампициллину, соответственно; lacI - ген репрессора LacI; Pspac - IPTG-зависимый промотор; t1t2t0, терминаторы(lt0; t1, t2 из rrnB), trpAt, терминатор триптофанового оперона E.coli; MCS, полилинкер; tag, последовательность, кодирующая антигенную детерминанту(FLAG, cMyc или HA); FP, ген флуоресцентного белка (GFP+, CFP или YFP).
На основе вектора pMUTIN2 (Vagneretal., 1998; см. выше) был сконструирован вектор pMUTIN-Ter (рис.12а). На его основе, путём вставки последовательностей, кодирующих соответствующие антигенные детерминанты, были получены векторы pMUTIN-FLAG, -cMyc и -HA. Для проведения слияния интересующего белка с этими антигенными детерминантами, 3¢-конец гена (около 300п.н.) вставляется непосредственно перед последовательностью, кодирующей антигенную детерминанту. Нужный гибридный белок будет синтезироваться после интеграции целого вектора в хромосомальный ген путём гомологичной рекомбинации. Если ген является частью оперона, то транскрипция проксимальнее расположенных генов оперона обеспечивается IPTG-зависимым промотором. Антигенные детерминанты c-Myc и HA могут быть также использованы для очистки гибридных белков путём аффинной хроматографии.
Аналогично на основе pMUTIN-Ter путём вставкипоследовательностей, кодирующих соответствующие флуорисцирующие белки, были получены векторы pMUTIN-GFP+, -CFP и –YFP (рис.12b), позволяющие производить слияние белков, кодируемых хромосомальными генами, с флуоресцирующими белками. Это позволяет проследить локализацию исследуемого белка в клетке. Функционирование всех шести векторов было протестировано на цитоплазматическом белке HtpG (белок теплового шока) и интегральном мембранном белке FtsH (протеаза).
1.7. Геномные векторы
Для клонирования больших участков генома на основе генома Bacillus subtilis168 создан геномный вектор - BGM вектор (BacillusGenoMe вектор) (Itayaetal., 2000; Itayaetal., 2003; Kanekoetal., 2003). В отличие от плазмидных векторов, в которые нужные фрагменты ДНК вставляются путём непосредственного лигирования, введение вставки в BGM вектор происходит путём гомологичной рекомбинации. Для этого BGM вектор должен иметь последовательности гомологичные участкам ДНК, фланкирующим клонируемый фрагмент. Данные последовательности, называемые LPS (Landing Pad Sequences) (Itaya etal., 2000), приготавливаются в плазмиде pBR322 (или её производных) и вставляются в pBR322 последовательность генома B.subtilisпутём трансформации и общей рекомбинации. LPS составляют 5-10% от клонируемой вставки. Они вставляются вместе с фланкируемым ими маркером, что позволяет в дальнейшем проводить отбор рекомбинантов. LPS вместе с маркером обозначены как LPA; соответственно плазмиды, с помощью которых осуществляется вставка LPA в геном (в BGM вектор), называются LPA плазмидами, а штаммы B.subtilis со вставкой LPA в геноме – LPA штаммами.Вставку клонируемого сегмента в BGM вектор производят путём трансформации соответствующего LPA штамма; клонируемый сегмент, заключённый между двумя LPS, интегрируется в хромосому путём гомологичной рекомбинации по обоим LPS.
С помощью данной методики были клонированы разнообразные фрагменты ДНК как прокариотического, так и эукариотического происхождения.
Так, было клонировано несколько фрагментов генома фотосинтетической бактерии Synechocystis (Itayaetal., 2003; рис.13). Эта бактерия была выбрана по двум причинам: известна нуклеотидная последовательность её генома и отсутствие патогенности.
В одной серии экспериментов для селекции использовался собственный маркер BGM вектора, позволяющий клонировать любой участок генома. В качестве BGM вектора выступал штамм BEST7003, несущий две вставки: 4,3-kb последовательности pBR322 в гене proB и ген устойчивости к неомицину pr-neo вставленный в ген yah. Интегрированная в хромосому последовательность pBR322 состоит из двух частей: 2.4-kb фрагмент включающий в себя генb-лактамазы (ген устойчивости к ампициллину) и 1.9-kb фрагмент, включающий ген устойчивости к тетрациклину. ДНК клонируется между этими двумя фрагментами как показано на рис.13. Экспрессия гена neo регулируется промотором Pr фагаλ, с которым способен специфично связываться репрессорный белок cI.

Рис.13. Клонирование генома Synechocystisв геномный вектор B.subtilis(BGM). BEST6016 и BEST7003, геномные векторы для прямой и непрямой селекции. Структура промежуточного генома показана в скобках. Х означает гомологичную рекомбинацию. Геномная последовательность pBR322 представлена жёлтым (часть, содержащая ген b-лактамазы) и синим (часть, содержащая ген устойчивости к тетрациклину) заштрихованными боксами разделёнными сайтом для клонирования. Гены резистентности обозначены заштрихованным кругом (хлорамфеникол),незаштрихованным кругом (эритромицин), заштрихованным треугольником (тетрациклин), заштрихованным ромбом (спектиномицин). Изогнутая стрелка обозначает супрессию промотора Pr продуктом гена CI. [I] означает сайт для рестриктазы I-PpoI.
В LPA плазмидах между двумя LPS находился ген cI, кодирующий репрессор CI, и ген устойчивости к спектиномицину; между тетрациклиновой частью pBR322 и правым LPS находился ген устойчивости к хлорамфениколу. LPA плазмиды были приготовлены в E.coliи затем трансформировались вBEST7003. В результате трансформанты (LPA штаммы) становились чувствительными к неомицину (репрессировался cI) и приобретали устойчивость к спектиномицину и хлорамфениколу (маркеры из LPA). Далее LPA штаммы трансформировались препаратом генома дикого штамма Synechocystis PCC6803. Вставка части генома Synechocystisзамещала фрагмент cI-spcR, в результате чего дерепрессировался neo и терялся маркер устойчивости к спектиномицину. Поэтому клоны устойчивые к неомицину и чувствительные к спектиномицину несли нужную вставку.
Таким образом, были клонированы три сегмента генома Synechocystis: A,B и C длиной 26-, 38-, 43-kb соответственно). Для каждого сегмента создавались свои LPS, LPA плазмиды и LPA штаммы. Попытки клонировать сегменты длиннее 50 kb оказались безуспешными.
Поэтому в другой серии экспериментов был использован метод прямой селекции (рис.13), который также был применён для количественной оценки зависимости эффективности клонирования от размера вставки. Из дикого штамма Synechocystis PCC6803 был получен мутантный штамм BUSY1001, который в гене rnhB содержит вставку гена устойчивости к спектиномицину. Данный маркер и использовался для проведения прямой селекции рекомбинантов со вставкой генома Synechocystis .
В качестве BGM вектора выступал штамм B.subtilis BEST6016, несущий последовательность pBR322, вставленная в ген proB(как и BEST7003), а также ген устойчивости к неомицину [Pr-neo], вставленный в ген yvfC(неомициновый маркер BEST6016 в данных экспериментах не использовался).
В LPA плазмидах между двумя LPS находился ген устойчивости к эритромицину, и LPA штаммы отбирали по этому маркеру. Таким образом, были созданы 4 штамма B.subtilis для клонирования сегментов, содержащих rnhB::spc, длиной от 14 до 77 kb. После трансформации препаратом генома Synechocystis BUSY1001 шёл прямой отбор рекомбинантов по их устойчивости к спектиномицину. Было показано, что эффективность клонирования понижается с увеличением размера клонируемого сегмента.
С помощью BGM вектора BEST6016 также был клонирован 120-kb фрагмент геномной ДНК мыши (Itayaetal.,2000). Клонирование фрагмента шло в несколько этапов (рис.14). Сначала вBEST6016 клонировали первый фрагмент длиной 20 kb, а затем путём трансформации и гомологичной рекомбинации к нему были добавлены несколько фрагментов, в результате чего он был удлинён до 120 kb. Все проведённыеманипуляции были аналогичны описанным выше.
С помощью BGM вектора BEST7003 был клонирован ген мыши jumonji(jmj) (Kanekoetal., 2003). Данный ген имеет размер около 200 kb. Предполагалось, что с первым интроном длиной 90 kbвзаимодействуют какие-то белковые факторы, благодаря чему происходит регуляция экспрессии данного гена. Чтобы проверить это, были получены фрагменты данного интрона разной длины. Для этого в векторBEST7003 аналогично описанной процедуре ввели 110 kb фрагмент гена, включающий первые два интрона. Далее с помощью гомологичной рекомбинации была произведена серия делеций (рис.15) и были получены несколько сегментов первого интрона различной длины. Процесс введения делеций по сути является обратным процессу удлинения фрагмента.
|
|
BEST2257 |
|
[1] |
|
[2] |
|
[3] |


Рис.14. Объединение двух
фрагментов ДНК мыши длиной по 25 kb в один фрагмент длиной 50 kb. S2, S1, M1 - LPS-последовательности. BEST2257, BGM
вектор BEST6016 со
вставкой первого 25 kb
фрагмента. neo,
ген устойчивости к неомицину, Показаны следующие этапы: [1] клонирование S1и S2 в плазмиде производной pBR322 (между S1и S2 расположен гены cI); [2] вставка в BEST2257 фрагментов S1и S2 вместе с геном cI
( neo продуктом гена cI). Вставка происходит в результате гомологичной рекомбинация по
LPS-последовательности S1 и части pBR322 последовательности; [3] замещение
cI вторым 25 kb фрагментом.
neo продуктом гена cI). Вставка происходит в результате гомологичной рекомбинация по
LPS-последовательности S1 и части pBR322 последовательности; [3] замещение
cI вторым 25 kb фрагментом.
Во всех вышеописанных экспериментах клонированные в геномном векторе сегменты реплицировались как часть генома B.subtilis, поэтому они проявляли высокую структурную и сегрегационную стабильность. Клонированная ДНК стабильно сохранялась в геноме даже при выращивании в неселективной среде. При этом скорость роста рекомбинантов со вставкой части генома была такой же, как и у клеток без вставки.
|
|
|
|

Рис.15. Осуществление делеции участка клонированного фрагмента. Вверху: схема клонированного фрагмента гена jmj(длина фрагмента 110kb). Белые прямоугольники, экзоны. Внизу: делеция участка длиной 24 kb путём гомологичной рекомбинации по двум LPS (внутренний участок вставки и участок последовательности pBR322).
Полученные результаты показывают, что в геномном векторе можно клонировать ДНК практически любой типа. Приготовление двух фланкирующих сегментов перед созданием BGM неизбежно, однако позволяет осуществлять позиционное клонирование молекул ДНК размерами, превышающими лимит технологии ПЦР. Клонирование с помощью геномного вектора имеет несколько преимуществ по сравнению с клонированием с помощью плазмидных векторов. Процесс клонирования в плазмидных векторах сильно зависит от рестрицирующих эндонуклеаз и наличия определённых сайтов рестрикции. Кроме того, при использовании таких векторов невозможны дальнейшие манипуляция с клонированной вставкой. Клонированная в геномном векторе ДНК не отличается от остальной геномной ДНК в отношении её способности участвовать в процессах рекомбинации. Это позволяет проводить с ней различные манипуляции: вставки, делеции, замещения и добавление примыкающих сегментов. Кроме того, одним из преимуществ является возможность хранения клонированной ДНК при комнатной температуре в спорах B.subtilis.
ЗАКЛЮЧЕНИЕ
К настоящему моменту для B.subtilis разработано большое количество как непосредственно клонирующих векторов, так и специализированных векторов: для клонирования промоторов и терминаторов, для экспрессии, получения гибридных белков, векторов с регулируемой копийностью и векторов для осуществления направленной инактивации генов.
В основе векторов для B.subtilis лежат плазмидные реликоны, выделенные из близкородственных организмов, таких какBacillus thuringiensis, Enterococcus faecalis,Staphylococcus aureus либо Lactococcus lactis.
Большинство из указанных векторов могут быть также использованы для ряда других грамположительных бактерий, т.к. репликоны, на основе которых они построены, часто могут обеспечивать репликацию и поддержание векторов в клетках бактерий принадлежащих к нескольким близким видам и даже родам.
.
ЛИТЕРАТУРА
1. Сельскохозяйственная биотехнология: векторные системы молекулярного клонирования / Под ред. В.И.Негрука.- М.: Агропромиздат, 1991.- 534 с.
2. Ballester S., Lopez P., Espinosa M., Alonso J.C, Lacks S.A. Plasmid structural instability associated with pC194 replication functions // J.Bacteriol. - 1989. – V.171. – p. 2271-2277.
3. Bhavsar A.P., Zhao X., Brown E.D. Development and characterization of a xylose-dependent system for expression of cloned genes in Bacillus subtilis: conditional complementation of a teichoic acid mutant //Applied and environmental microbiology.- Jan.2001.-p.403-410.
4. Cutting, S. M., and P. B. V. Horn. 1990. Genetic analysis, p. 27–61. In C. R. Harwood and S. M. Cutting (ed.), Molecular biological methods for Bacillus. John Wiley and Sons, Toronto, Canada.
5 . Dabert P., Ehrliсh S.D., Gruss A. High-molecular-weight linear multimer formation by single stranded DNA plasmids in Escherichia coli // J. Bacteriol. - 1992. – V.174. – p.173-178.
6 . Dabert P., Ehrliсh S.D., Gruss A. CHI sequence protects against RecBCD degradation of DNA in vivo // Proc. Natl. Acad. Sci. USA. - 1992. – V.89. – p.12073-12077.
7. Duffner F., Bertoldo C., Andersen J.T., Wagner K., Antrakian G. A new thermoactive pullulanase from Desulfurococcus mucosus: cloning, sequencing, purification, and characterization of the recombinant enzyme after expression in Bacillus subtilis. // J.Bacteriol. -2000.-V.182.- p.6331-6338.
8. Ehrlich S.D., Noirot P., Petit M.A., Janniere L., Michel B., te Riele H. Structural instability of Bacillus subtilis plasmids. // In Setlow, J.K. and Hollaender, A. (Eds.), Genetic Engineering, Vol.8. Plenum, New York, 1986, pp. 71-83.
9. Feucht A., Lewis P.J.Improved plasmid vectors for the production of multiple fluorescent protein fusions in Bacillus subtilis // Gene.- 2001.-V.264.- p.289-297
10. Gros M. -F., te Riele H., Ehrlich S.D. Replication origin of a single-stranded DNA plasmid pC194 // EMBO J. - 1989. – V.8. – p.2711-2716.
11. Gruss A., Ehrlich S.D. Insertion of foreign DNA into plasmids from gram-positive bacteria induces formation of high-molecular-weight plasmid multimers // J. Bacteriol. - 1988. – V.170. – p.1183-1190.
12. Itaya, M., Shiroishi, T., Nagata, T., Fujita, K., and Tsuge, KEfficient cloning and engineering of giant DNAs in a novel Bacillus subtilis genome vector // J. Biochem.- 2000- V.128.- p.869-875.
13. Itaya M., Fujita K., Ikeuchi M., Koizumi1M., Tsuge K. Stable positional cloning of long continuous DNA in the Bacillus subtilis genome vector // J. Biochem.- 2003.- V.134.- p.513-519.
14. Janniere L., Bruand C., Ehrlich S.D. Structurally stable Bacillus subtilis cloning vectors // Gene. - 1990. – V.87. - p.53-61.
15. Kaltwasser M., Wiegert T.,Schumann W. Construction and application of epitope- and green fluorescent protein-tagging integration vectors for Bacillus subtilis// Applied and environmental microbiology.-2002.- V. 68.-No.5.- p.2624-2628.
16. Kaneko S., Tsuge K., Takeuchi T., Itaya M. Conversion of sub-megasized DNA to desired structures using a novel Bacillus subtilis genome vector // Nucleic Acids Research.-2003.- V. 31.- No.18.- e112
17. Lam K.H.E., Chow K.C., Wong. Construction of an efficient Bacillus subtilis system for extracellular production of heterologous proteins. // Journal of Biotechnology.-1998.-V.63.- p.167-177.
18. Leonard С, Zekri O, Mahillon J. Integrated Physical and Genetic Mapping of Bacillus cereus and other Gram-Positive Bacteria Based on IS231A transposition Vectors // Infection and Immunity.-1998. - V.66.- No.5. - p.2163-2169.
19. Lewis P.J, Marston A.L. GFP vectors for controlled expression and dual labeling of protein fusions in Bacillus subtilis // Gene.- 1999.- V.227.- p.101–109
20. Michel B., Palla E., Niaudet B., Ehrlich S.D. DNA cloning in Bacillus subtilis, III. Efficiency of random-segment cloning and insertional inactivation vectors. // Gene. –1980. –V. 12. – p.147-154.
21. Michel B., Ehrlich S.D. Illegitimate recombination occurs between the replication origin of plasmid pC194 and a progressive replication fork. // EMBO J. - 1986. – V.5. – p.3691-1696.
22. Poyart C., Tieu-Cuot P. A broad-host-range mobilizable shuttle vector for the construction of transcriptional fusion to b-galactosidase in Gram-positive bacteria. // FEMS Microbiology Letters. – 1997. – V.156. – p.193-198.
23. Renault P., Cothier G., Goupil N., Delorme C., Ehrliсh D.S. Plasmid vectors for Gram-positive bacteria switching from high to low copy number // Gene. – 1996. – V.183. – p.175-182.
24. Takao M., Yamaguchi A., Yoshikawa K., Terashita T., Sakai T. Molecular cloning of the gene encoding thermostable endo-1,5-a-L-arabinase of Bacillus thermodenitrificans TS-3 and its expression in Bacillus subtilis // Biosci. Biotechnol. Biochem. – 2002.- V.66(2).-p.430-433.
25. Weickert, M. J., and G. H. Chambliss. Site-directed mutagenesis of a catabolite repression operator sequence in Bacillus subtilis // Proc. Natl. Acad. Sci. USA.- 1990.- V.87.- p.6238–6242.
26. Vagner V., Dervyn E., Ehrlich S.D. A vector for systematic gene inactivation in Bacillus subtilis. // Microbiology. – 1998. – V.144. – p.3097-3104.